Akademia Wychowania Fizycznego im. Eugeniusza Piaseckiego w Poznaniu, Poznań 2003.
Spis treści
- 1. Wstęp*
- 2. Uzasadnienie wyboru testowych zadań motorycznych do badań własnych
- 3. Cele i hipotezy
- 4. Materiał badań
- 5. Metody badawcze
- 5.1. Pomiar siły i dokładności siłowej
- 5.2. Pomiar prędkości biegowej
- 5.3. Pomiar szybkości reakcji i dokładności czasowej
- 5.4. Pomiar prędkości pojedynczych ruchów
- 5.5. Pomiar wysokości wyskoku pionowego
- 5.6. Pomiar maksymalnego zakresu obrotu w wyskoku
- 5.7. Pomiar częstotliwości ruchów globalnych
- 5.8. Pomiar częstotliwości ruchów lokalnych
- 5.9. Pomiar zdolności zachowania równowagi statycznej
- 5.10. Ocena dokładności przejawiania zdolności motorycznych
- 5.11. Metody statystyczne
- 5.12. Rzetelność stosowanych testowych zadań motorycznych
- 6. Wyniki badań
- 6.1. Zależności pomiędzy kondycyjnymi zdolnościami motorycznymi
- 6.2. Zależności pomiędzy koordynacyjnymi zdolnościami motorycznymi
- 6.3. Zależności pomiędzy kondycyjnymi a koordynacyjnymi zdolnościami motorycznymi
- 6.4. Lokalne a globalne zdolności motoryczne
- 6.5. Kondycyjne a koordynacyjne zdolności motoryczne
- 6.6. Charakterystyka podobieństw pomiędzy badanymi zdolnościami motorycznymi
- 6.7. Maksymalne i optymalne przejawy zdolności motorycznych o globalnym i lokalnym charakterze
- 6.8. Czas narastania siły w zadaniach o maksymalnym i optymalnym charakterze jako wyraz specyficznych wymogów związanych z dokładnością siłową
- 6.9. Zdolność do dokładnego odtwarzania zadanej wartości zdolności motorycznych a rodzaj ruchu
- 6.10. Symetryczność globalnych i lokalnych przejawów kondycyjnych oraz koordynacyjnych zdolności motorycznych
- 6.11. Zróżnicowanie płciowe globalnych i lokalnych przejawów kondycyjnych i koordynacyjnych zdolności motorycznych
- 7. Dyskusja
- 7.1. Związki pomiędzy zdolnościami motorycznymi
- 7.2. Globalne a lokalne zdolności motoryczne
- 7.3. Kondycyjne a koordynacyjne zdolności motoryczne z uwzględnieniem ich globalnych i lokalnych przejawów
- 7.4. Maksymalne i optymalne przejawy globalnych i lokalnych zdolności motorycznych
- 7.5. Symetria w zakresie przejawiania globalnych i lokalnych zdolności motorycznych
- 7.6. Zróżnicowanie płciowe w zakresie globalnych i lokalnych przejawów zdolności motorycznych
- 8. Wnioski
- Piśmiennictwo
- Summary
- Zusammenfassung
Spis rysunków
- Rycina 1 Struktura zdolności motorycznych człowieka [Gundlach 1970]
- Rycina 2 Koncepcja poziomów koordynacji ruchowej Farfela [Starosta 1987]
- Rycina 3 Podstawowe zdolności koordynacyjne i ich powiązania w kompleksy koordynacyjne [Blume 1981]
- Rycina 4 Koncepcja hierarchicznej struktury podstawowych zdolności koordynacyjnych [Hirtz 1985]
- Rycina 5 Schemat hierarchicznego ujęcia podstawowych zdolności motorycznych [Starosta 1995a]
- Rycina 6 Schemat współdziałania analizatorów przy tworzeniu „czucia ruchu” [Hotz i Weineck 1983]
- Rycina 7 Grupy skupień zdolności motorycznych (n = 176)
- Rycina 8 Siła i czas jej narastania w pomiarach maksymalnej globalnej siły statycznej i globalnej dokładności siłowej (n = 176)
- Rycina 9 Siła i czas jej narastania w pomiarach maksymalnej lokalnej siły statycznej i lokalnej dokładności siłowej (n = 176)
- Rycina 10 Charakterystyki siły statycznej i dokładności siłowej (n = 176)
- Rycina 11 Zestawienie średnich wartości błędów popełnianych w zadaniach motorycznych wymagających przejawienia największej dokładności (n = 176)
- Rycina 12 Względne wartości symetrycznych przejawów motoryczności oraz kształtowanie się wskaźnika symetryzacji (n = 176)
- Rycina 13 Względne wartości zdolności motorycznych w zespołach kobiet (n = 84) i mężczyzn (n = 176) – kondycyjne zdolności motoryczne
- Rycina 14 Względne wartości zdolności motorycznych w zespołach kobiet (n = 84) i mężczyzn (n = 176) - koordynacyjne zdolności motoryczne
- Rycina 15 Istotne zależności pomiędzy globalnymi a lokalnymi zdolnościami motorycznymi (n = 176)
- Rycina 16 Zależności pomiędzy globalnymi i lokalnymi przejawami kondycyjnych i koordynacyjnych zdolności motorycznych (n = 176)
- Rycina 17 Zestawienie wskaźników symetrycznego przejawiania zdolności moto-rycznych w zespole badanych studentów (n = 176)
- Rycina 18 Zestawienie wskaźników symetryzacji w parach zadań motorycznych wymagających maksymalnej i optymalnej aktywności (n = 176)
- Rycina 19 Błędy w zadaniach motorycznych wymagających maksymalnej dokładności (mężczyźni n = 176; kobiety n = 84)
- Rycina 20 Zestawienie wskaźników dymorfizmu kondycyjnych i koordynacyjnych zdolności motorycznych (mężczyźni n = 176; kobiety n = 84)
Spis tabel
- Tabela 1 Zestawienie zastosowanych w pracy zadań motorycznych na podstawie klasyfikacji podstawowych zdolności koordynacyjnych [Raczek i wsp. 2002; Waśkiewicz 2002]
- Tabela 2 Zestawienie zastosowanych w pracy zadań motorycznych w odniesieniu do klasyfikacji dzielącej zdolności motoryczne na przejawy szybkiej i wolnej koordynacji [Hirtz 1985]
- Tabela 3 Masa i wysokość ciała badanych studentów i studentek
- Tabela 4 Porównanie istotności różnic pomiędzy średnimi arytmetycznymi w obrębie dwóch grup złożonych ze studentów o najniższej (n = 30) i najwyższej (n = 30) wysokości ciała
- Tabela 5 Porównanie istotności różnic pomiędzy średnimi arytmetycznymi w obrębie dwóch grup złożonych ze studentów o najmniejszej (n = 30) i największej (n = 30) masie ciała
- Tabela 6 Zestawienie istotności różnic pomiędzy średnimi wartościami zdolności motorycznych w grupach złożonych z badanych mężczyzn o największym (n = 30) i najmniejszym (n = 30) BMI
- Tabela 7 Zestawienie uwzględnionych w badaniach globalnych i lokalnych kondycyjnych zdolności motorycznych
- Tabela 8 Zestawienie uwzględnionych w badaniach globalnych i lokalnych koordynacyjnych zdolności motorycznych
- Tabela 9 Zestawienie kondycyjnych i koordynacyjnych zdolności motorycznych o globalnym oraz lokalnym charakterze
- Tabela 10 Zestawienie zadań ruchowych zastosowanych do analizy maksymalnych i optymalnych przejawów zdolności motorycznych
- Tabela 11 Miary rzetelności zadań motorycznych zastosowanych w badaniach własnych
- Tabela 12 Średnie wartości kondycyjnych zdolności motorycznych (n = 176)
- Tabela 13 Statystycznie istotne współczynniki korelacji w obrębie globalnych kondycyjnych zdolności motorycznych (n = 176)
- Tabela 14 Statystycznie istotne współczynniki korelacji w obrębie lokalnych kondycyjnych zdolności motorycznych (n = 176)
- Tabela 15 Statystycznie istotne współczynniki korelacji pomiędzy globalnymi a lokalnymi kondycyjnymi zdolnościami motorycznymi (n = 176)
- Tabela 16 Średnie wartości koordynacyjnych zdolności motorycznych (n = 176)
- Tabela 17 Statystycznie istotne współczynniki korelacji w obrębie globalnych koordynacyjnych zdolności motorycznych (n = 176)
- Tabela 18 Statystycznie istotne współczynniki korelacji w obrębie lokalnych koordynacyjnych zdolności motorycznych (n = 176)
- Tabela 19 Statystycznie istotne współczynniki korelacji pomiędzy globalnymi a lokalnymi koordynacyjnymi zdolnościami motorycznymi (n = 176)
- Tabela 20 Zestawienie statystycznie istotnych współczynników korelacji pomiędzy kondycyjnymi a koordynacyjnymi zdolnościami motorycznymi (n = 176)
- Tabela 21 Charakterystyki statystycznie istotnych równań regresji wielokrotnej pozostałe po redukcji zmiennych niezależnych. Globalne a lokalne zdolności kondycyjne (n = 176)
- Tabela 22 Charakterystyki statystycznie istotnych równań regresji wielokrotnej pozostałe po dokonaniu redukcji zmiennych niezależnych. Kondycyjne a koordynacyjne zdolności motoryczne (n = 176)
- Tabela 23 Zestawienie współczynników korelacji w parach zdolności motorycznych (maksymalne – optymalne) (n = 176)
- Tabela 24 Charakterystyki statystycznie istotnych równań regresji wielokrotnej pomiędzy zdolnościami motorycznymi o maksymalnym i optymalnym charakterze aktywności (n = 176)
- Tabela 25 Lokalne uwarunkowania globalnych przejawów zdolności motorycznych – maksymalne (n = 176)
- Tabela 26 Maksymalne uwarunkowania optymalnych przejawów zdolności motorycznych – globalne (n = 176)
- Tabela 27 Maksymalne uwarunkowania optymalnych przejawów zdolności motorycznych – lokalne (n = 176)
- Tabela 28 Zestawienie istotności różnic pomiędzy średnimi wartościami błędów w zadaniach optymalizacyjnych (n = 176)
- Tabela 29 Charakterystyki symetrycznych zdolności motorycznych (n = 176)
- Tabela 30 Istotności różnic w kondycyjnych i koordynacyjnych zdolnościach motorycznych u mężczyzn (n = 176) i kobiet (n = 84)
- Tabela 31 Zestawienie wartości błędów popełnianych przez mężczyzn (n = 176) i kobiety (n = 84) w zadaniach motorycznych wymagających największej dokładności
Wiedza o motoryczności człowieka jest podstawowym zagadnieniem w naukach o kulturze fizycznej. Za jednego z twórców współczesnej teorii motoryczności uznaje się Bernsteina [1947], który w swojej pracy „O strukturze ruchów” zawarł jej podstawy. Zgodnie z nią działanie ruchowe człowieka polega na nadążającym porównywaniu pożądanej wartości z aktualną, dotyczącą wskaźników charakteryzujących ruch. Rozpoczęcie ruchu jest wg Bernsteina możliwe po wyobrażeniu celu i skonstruowaniu programu działania. Zatem koordynacja ruchowa to „(...) pokonywanie nadmiernej liczby stopni swobody poruszającego się organizmu, czyli przekształcenie go w system sterowalny” [Bernstein 1966]. Do dalszych rozważań przyjęto definicję koordynacji ruchowej jako zdolności „(...) do wykonywania złożonych ruchów dokładnie, szybko i w zmiennych warunkach” [Szopa 1989a; Starosta 1984].
Natomiast Meinel w monografii „Bewegungslehre” wskazał czynność motoryczną jako przedmiot teorii motoryczności [Meinel 1967]. Twierdził on, że należy ruchy analizować w ich zewnętrznych przejawach tak, aby ujawniać i poznawać podstawowe mechanizmy oraz procesy wewnętrzne, determinujące skuteczność ruchu.
Gundlach [1968, 1970] dokonał klasyfikacji zdolności motorycznych opartej na podłożu funkcjonalnym. Wyróżnił on zdolności kondycyjne zdeterminowane mechanizmami energetycznymi i zdolności koordynacyjne uwarunkowane w zasadniczym stopniu procesami sterowania, regulacji i kognicji (ryc. 1).
Ta koncepcja, pomimo późniejszych modyfikacji, stała się podstawową w analizowaniu struktury zdolności motorycznych [Raczek 1987; Hirtz, Starosta 1990; Starosta 1995a]. Raczek [1991a] dodatkowo wyodrębnił grupę zdolności hybrydowych, pośrednią pomiędzy kondycyjnymi a koordynacyjnymi zdolnościami motorycznymi.
Bardzo szerokie spektrum uwarunkowań przejawiania się zdolności motorycznych powoduje, że brak jest definicji akceptowanych przez większość badaczy. Wielu z nich nawołuje do stworzenia jasnej koncepcji motoryczności człowieka [Meinel 1967; Zaciorski 1970; Fidelus 1972; Raczek 1986; Osiński 1990; Hirtz, Starosta 1990; Szopa 1993]. Dlatego problem klasyfikacji zdolności motorycznych można ciągle uznawać za otwarty.
W pracy tej używa się pojęcia „zdolność motoryczna” a nie „cecha”. Założono też, że poszczególne zdolności motoryczne przejawiają się w odpowiednio dobranych zadaniach ruchowych. Raczek [1987] i Szopa [1986] wskazywali na trudności w określeniu rzeczywistych cech motoryczności człowieka. Powoduje to często rozbieżności pomiędzy stosowanymi definicjami a używanymi metodami badawczymi. „Cechy” zależą od uwarunkowań genetycznych, natomiast „zdolności”, pomimo znacznego wrodzonego zdeterminowania, mogą być modyfikowane pod wpływem czynników środowiskowych. Sankowski [1990] uważa, że koncepcję „cech motorycznych” od teorii „zdolności motorycznych” różni głównie udział czynników nabytych w przejawianiu się zdolności motorycznych. Także wielu autorów stosujących określenie „cechy motoryczne” swoje definicje rozpoczyna od stwierdzenia, że są to zdolności [Denisiuk, Milicerowa 1969; Zaciorski 1970; Ważny 1977; Naglak 1979].
![Rycina 1 Struktura zdolności motorycznych człowieka [Gundlach 1970]](rysunki/image002.gif)
Najczęściej badano zagadnienia związane z kondycyjnymi zdolnościami motorycznymi. Większość autorów uznało siłę, wytrzymałość i szybkość za podstawowe zdolności kondycyjne. Jedynie Raczek [1993] klasyfikuje tę ostatnią zdolność w grupie zdolności hybrydowych – pośrednich. Problematyce kondycyjnych zdolności motorycznych poświęcono szereg opracowań [Farfel 1960; Meinel 1967; Morecki i wsp. 1971; Měkota 1972; Schnabel 1974; Wolański, Pařizkowa 1976; Schmidt 1976, 1988; Celikovsky i wsp. 1978; Kasa 1983; Starosta 1984, 1987, 1993a; Rutkowska-Kucharska, Bober 1986; Blume 1981; Hirtz 1985, 1989; Meinel, Schnabel 1998; Raczek 1987, 1991b, 1993; Osiński 1977, 1988a; Szopa 1989b, 1992; Raczek, Mynarski 1991, 1992; Szopa i wsp. 1999; Wachowski, Strzelczyk 1999].
Wszechstronnym badaniom poddano też podstawowe przejawy koordynacyjnych zdolności motorycznych [Farfel 1960; Meinel 1967; Morecki, i wsp. 1971; Měkota 1972; Schnabel 1974; Wolański, Pařizkowa 1976; Celikovsky i wsp. 1979; Kasa 1983; Starosta 1984, 1987, 1993a; Rutkowska-Kucharska, Bober 1986; Blume 1981; Hirtz 1985, 1989; Meinel, Schnabel 1998; Raczek 1987, 1991b, 1993; Osiński 1981, 1988b; Szopa 1989a, 1992; Raczek, Mynarski 1991, 1992; Szopa i wsp. 1999; Mynarski 2000; Ljach 2001; Raczek i wsp. 2002; Waśkiewicz 2002]. Trudności napotykało natomiast precyzyjne zdefiniowanie koordynacji ruchowej, dlatego spotyka się szereg rozmaitych definicji [Gilewicz 1964; Szopa 1988; Raczek, Mynarski 1991; Szopa 1989a; Starosta 1987]. Znaczenie koordynacji ruchowej jest szczególnie duże, gdyż stanowi ona bazę rozwoju techniki ruchu i jej przejawiania w konkretnych zadaniach ruchowych. Dlatego Hirtz [1989] używa w stosunku do niej określenia „fenomenalna”. Natomiast Starosta [1990] określił ją jako „super cechę”. Nie można rozpatrywać koordynacji ruchowej w oderwaniu od analizy funkcjonowania centralnego układu nerwowego. Koordynacja ruchowa zależy bowiem od działania receptorów, wydzielania i transportu przez krew hormonów oraz od przepływu informacji przez układ nerwowy. Dokładne, zatem skoordynowane wykonywanie ruchów wymaga stworzenia programów działania przed ich rozpoczęciem. W trakcie ruchu następuje systematyczne ocenianie jego przebiegu i dokonywanie korekt na podstawie rozbieżności pomiędzy osiąganym rezultatem a zaplanowanym celem działania [Bernstein 1975; Schnabel 1968, 1974; Starosta 1993a; Blume 1978a, 1978b, 1981; Hirtz 1989; Mattausch 1973; Pöhlmann i wsp. 1979; Zimmermann i Nicklisch 1981].
Stopień złożoności koordynacji ruchowej obrazuje ułożenie zdolności koordynacyjnych (rozumianych jako rozmaite specyficzne przejawy koordynacji ruchowej) według ich hierarchii. Farfel [1960] podzielił je na trzy poziomy o różnym stopniu złożoności. Koncepcję tę zilustrował i poszerzył Starosta [1987] (ryc. 2).
![Rycina 2 Koncepcja poziomów koordynacji ruchowej Farfela [Starosta 1987]](rysunki/image004.gif)
I poziom – charakteryzuje przestrzenną dokładność czynności ruchowych wykonywanych w standardowych warunkach bez ograniczenia czasowego.
II poziom – wyznacza przestrzenną dokładność ruchu w optymalnym – ograniczonym czasie w standardowych warunkach.
III poziom – charakteryzuje zadania ruchowe wykonywane dokładnie, szybko lub z dostosowaniem szybkości do zmiennych warunków zewnętrznych.
Zasadność tej koncepcji potwierdziły inne badania [Rutkowska-Kucharska, Bober 1986; Starosta 1993a].
Powstało też wiele innych teorii. Celikovsky i wsp. [1979] także wydzielili trzy poziomy koordynacji. Na najniższym umieszczono zdolności różnicowania kinestetycznego, równowagi, rytmizacji oraz orientacji przestrzennej. Są one związane z podstawowymi właściwościami regulatora. Drugi poziom zawiera możliwości regulacji układu ruchu. Natomiast na trzecim, najwyższym poziomie umieszczono zdolność do oceny przestrzennej i czasowej struktury ruchów. Nazwano go poziomem regulacji czynności ruchowej, uznając za syntezę poprzednich. Stwierdzono, że warunkuje właściwe możliwości koordynacyjne, określone sprawnością układu nerwowego i elementów jego struktury [Celikovsky i wsp. 1979].
Również Blume [1981] wszystkie podstawowe zdolności koordynacyjne podzielił na trzy powiązane strukturalnie kompleksy. Są to: uczenie się motoryczne, sterowanie i regulacja ruchami oraz adaptacja motoryczna. Blume wyróżnia siedem podstawowych zdolności koordynacyjnych. Wszystkie są związane z kompleksem zdolności uczenia się motorycznego. Natomiast zdolności niezbędne do wykonywania ruchów standardowych (zdolność łączenia ruchów, różnicowania, równowagi, orientacji i rytmizacji) wiąże z kompleksem sterowania ruchami. W zespole zdolności warunkujących adaptację motoryczną umieścił te, które determinują możliwość dostosowania czynności ruchowych do zmiennych warunków i sytuacji. Zaliczył do nich zdolność orientacji, rytmizacji, reakcji i dostosowania motorycznego (ryc. 3).
![Rycina 3 Podstawowe zdolności koordynacyjne i ich powiązania w kompleksy koordynacyjne [Blume 1981]](rysunki/image006.gif)
W hierarchicznej koncepcji Hirtza [1985] także występuje kilka poziomów koordynacji ruchowej. Podstawową rolę w jej przejawianiu się odgrywają zdolności orientacji przestrzennej i kinestetycznego różnicowania ruchu, oparte na informacjach sensorycznych. Na kolejnym poziomie umieszczono bardziej kompleksowe funkcje ruchowe, których podłożem są złożone funkcje sensoryczne oraz skomplikowane programy ruchowe. Do typowych dla tego poziomu zdolności motorycznych Hirtz zaliczył zdolność szybkiej reakcji, rytmizacji i równowagi. Następny poziom związany jest z czasem realizacji zadania. W przypadku ograniczenia czasowego Hirtz używa określenia koordynacja szybka. Natomiast gdy ruch ma być wykonany z dużą dokładnością, jest to koordynacja wolna. Na najwyższym poziomie zdolności koordynacyjnych umieścił kompleksowe zdolności uczenia się motorycznego, zdolność sterowania ruchem i zdolność adaptacji motorycznej (ryc. 4).
![Rycina 4 Koncepcja hierarchicznej struktury podstawowych zdolności koordynacyjnych [Hirtz 1985]](rysunki/image008.gif)
Starosta [1993a] uważa, że pomimo istnienia trzech poziomów koordynacji ruchowej poszczególne zdolności są zintegrowane przez układ nerwowy i przejawiają się w tzw. „czuciach”. Na przykład „czucie wody” w pływaniu czy też różnego rodzaju czucia (piłki, sprzętu, powietrza, przeciwnika) występujące w innych dyscyplinach. Są one przejawem adaptacji do wykonywania określonych zadań ruchowych i wymagają odpowiednio rozwiniętych i właściwie ukierunkowanych wrażeń kinestetycznych. Starosta [1995a] zaproponował hierarchiczne ujęcie zdolności motorycznych, gdzie koordynacja ruchowa jest zdolnością integrującą i organizującą współdziałanie pozostałych zdolności motorycznych podczas wykonywania różnych zadań ruchowych (ryc. 5).
![Rycina 5 Schemat hierarchicznego ujęcia podstawowych zdolności motorycznych [Starosta 1995a]](rysunki/image010.gif)
Większość autorów zajmujących się problemami koordynacji ruchowej określa zdolność kinestetycznego różnicowania ruchu jako główną. Według Blume [1981] i Harre [1985] jest ona zdeterminowana umiejętnością dokładnego i ekonomicznego wykonania pojedynczych ruchów części ciała i poszczególnych faz cyklu ruchowego. Z jednej strony jest uwarunkowana świadomym i precyzyjnym określeniem parametrów siły, czasu i przestrzeni aktualnie wykonywanego ruchu oraz porównaniem z jego wyobrażonym wzorcem. Natomiast z drugiej strony jest uzależniona od dokładnej koordynacji ruchów stopy, dłoni i głowy oraz zdolności rozluźniania mięśni a także umiejętności świadomej regulacji ich napięcia. Przejawia się jako umiejętność dostosowania się do środowiska zewnętrznego. Zatem jest uznawana za podstawową zdolność warunkującą mistrzostwo sportowe.
Raczek i Mynarski [1992], dokonując syntezy opracowań innych autorów [Blume 1981; Harre 1985; Hirtz 1985], zastosowali termin „zdolność różnicowania kinestetycznego”. Uznali ją za warunkującą dokładne i ekonomiczne wykonywanie ruchów. O jej poziomie decydują trzy składowe, określane jako komponenty przestrzenne, czasowe i siłowe. Zawierają one odpowiednio informacje o kątowych pozycjach w stawach, napięciu pracujących mięśni oraz szybkości ruchów. Optymalny przebieg czynności ruchowej zapewnia odpowiednie dozowanie impulsów sterujących, poprzez precyzyjne postrzeganie siły czasu i przestrzeni podczas realizacji zadania ruchowego. W sterowaniu przebiegiem ruchu, tak aby był on dokładny i umożliwiał uzyskanie zaplanowanego efektu, najważniejszą rolę odgrywa zmysł kinestetyczny. Jest on decydujący w powstawaniu „czucia ruchu” [Hotz, Weineck 1983] (ryc. 6).
![Rycina 6 Schemat współdziałania analizatorów przy tworzeniu „czucia ruchu” [Hotz i Weineck 1983]](rysunki/image012.gif)
Dość często badano uwarunkowania poziomu zdolności motorycznych w zależności od wielu zmiennych. Zajmowano się jej zmianami w ontogenezie, ze szczególnym zwróceniem uwagi na okresy sensytywne i krytyczne. Mierzono jej zmiany u dzieci w zależności od wieku kalendarzowego i rozwojowego. Ze zrozumiałych względów częściej śledzono jej zmienność u dzieci niż u osób dorosłych [Starosta 1975, 1990, 1993a; Drabik 1983; Ljach 1989, 1990; Starosta, Hirtz 1989].
Wiele prac poświęcono określeniu wpływu na zdolności motoryczne treningu sportowego, prowadzonego przez zawodników o rozmaitym poziomie zaawansowania sportowego w różnych dyscyplinach sportowych. Dowodzono istnienia związku między środowiskiem społecznym a zdolnościami motorycznymi [Denisiuk, Milicerowa 1969; Osiński 1988a; Strzelczyk 1995].
Poszukiwano wpływu zmęczenia wywołanego wysiłkiem fizycznym na przejawianie się motoryki. Liczne prace ukierunkowano na określenie ewolucyjnych zmian ludzkiej motoryczności. Próbowano też ustalić związki występujące między rozwojem zdolności motorycznych a wykonywaną pracą zawodową. To ostatnie zagadnienie nabiera obecnie szczególnego znaczenia z powodu konieczności dostosowania się do specyfiki różnych zawodów. Coraz częściej nowe technologie powodują większe potrzeby związane z precyzyjnym wykonywaniem ruchów, kosztem typowych wymagań o większym podłożu energetycznym. Ponadto, w życiu człowieka konieczne staje się zmienianie profesji [Drozdowski 1981; Raczek i wsp. 2002].
Także problematyka genetycznych uwarunkowań koordynacyjnych zdolności motorycznych stała się przedmiotem coraz bardziej wszechstronnych badań [Rutkowska- -Kucharska 1982; Drozdowski Z. 1981; Mynarski 1986; Ljach 1990; Mleczko 1992, 1993; Szopa 1993; Starosta 2001a]. Jednak nie uzyskano jednoznacznych informacji o sile i rodzaju wrodzonego zdeterminowania koordynacyjnych zdolności motorycznych. Wielu autorów dowodzi znacznego stopnia ich genetycznego uwarunkowania [Balsewicz 1969, 2000; Skład 1973; Wolański, Kasprzak 1979; Ljach 1990]. Natomiast Szopa, Mleczko, Cempla [1985] oraz Mleczko [1991] twierdzą, że koordynacyjne zdolności motoryczne są najbardziej zmienne pod względem uwarunkowań genetycznych. Niezbyt wysoki poziom dziedziczności koordynacyjnych zdolności motorycznych sugeruje możliwość poprawy ich poziomu. Istnieje wiele dowodów skuteczności oddziaływania treningowego na koordynacyjne zdolności motoryczne [Zimmermann 1983, 1986; Szczepanik 1993; Szopa 1993; Szopa i wsp. 1996; Prus, Szopa 1997; Prus 2000]. Niemniej jednak konieczne jest ich uwzględnianie w procesie doboru do sportu oraz w ocenie przydatności do wykonywania niektórych zawodów [Raczek i wsp. 2002].
Natomiast istnieje niewiele opracowań dotyczących wzajemnych powiązań pomiędzy wskaźnikami budowy ciała i kondycyjnymi zdolnościami motorycznymi a koordynacyjnymi zdolnościami motorycznymi. W dotychczasowych badaniach stwierdzono, że koordynacyjne zdolności motoryczne niezbyt silnie były zdeterminowane wskaźnikami rozwoju somatycznego [Osiński 1988a; Ljach 1990; Strzelczyk 1995].
Wyjątkowo nieliczne publikacje dotyczyły powiązań pomiędzy lokalnymi a globalnymi przejawami zdolności motorycznych. Za ruchy lokalne uznano te, w których pracują mięśnie odpowiedzialne za ruch jednego stawu, z jednym stopniem swobody lub ruchy wykonywane całą kończyną z udziałem pewnej liczby stawów i mięśni, które nią poruszają. Natomiast ruchy wymagające zaangażowania większej liczby mięśni oraz odbywające się przy większej liczbie stopni swobody klasyfikowano jako globalne [Rutkowska-Kucharska 1982; Rutkowska-Kucharska i wsp. 1990]. W praktyce zawodowej i w sporcie często konieczne jest przejawianie rozmaitych zdolności w sposób globalny – angażujący większość grup mięśniowych i znacznie obciążających potencjał energetyczny zawodnika. Równocześnie istnieje jednak ogromna liczba zadań ruchowych, w których występuje jedynie lokalne zaangażowanie, tj. części organizmu.
W piśmiennictwie spotyka się określenia „mała motoryka” (fein Motorik) oraz „duża motoryka” (gross Motorik), jako odpowiedniki podziału ruchów na globalne i lokalne. Pierwsze określa precyzyjne ruchy części ciała, a drugie ruchy całego ciała [Mynarski 2000]. Mĕkota [1982] z czterech podstawowych zdolności koordynacyjnych wydzielił jedną o charakterze lokalnym. Nazwał ją zręcznością, rozumianą jako zwinność rąk. Ljach [1989, 1990], posługując się metodą analizy czynnikowej podzielił wszystkie zdolności koordynacyjne na dwie grupy. Do pierwszej zaliczył czynniki związane z wykonywaniem ruchów całym ciałem, a do drugiej – z ruchami rąk. Natomiast Starosta [2001a] sklasyfikował wszystkie ruchy jako globalne lub lokalne. Wyodrębnił siedem grup, z których w dwóch pierwszych umieścił ruchy lokalne (ruchy elementów ciała), a w pozostałych globalne (ruchy całego ciała). Pomimo znaczenia problematyki badań nad lokalnymi i globalnymi przejawami zdolności motorycznych w piśmiennictwie napotkano niewiele opracowań, zwłaszcza zawierających dane eksperymentalne [Rutkowska-Kucharska 1982; Rutkowska-Kucharska i wsp. 1990; Ljach 1989, 1990, 2001; Starosta 2001a].
W teorii treningu sportowego często wyznaczano cele i sposoby rozwijania siły z podziałem na jej globalne i lokalne przejawy [Zaciorski 1970; Trzaskoma, Trzaskoma 2001; Waśkiewicz 2002].
Przedstawione przykłady przeprowadzonych badań nad podziałem zdolności motorycznych na globalne i lokalne nie świadczą o dobrym rozpoznaniu tego problemu, a raczej o jego dostrzeżeniu. Dlatego podjęto próby dokonania analizy powiązań pomiędzy lokalnymi a globalnymi przejawami kondycyjnych i koordynacyjnych zdolności motorycznych. Są to zagadnienia związane z niezbyt często podejmowanym, ale ważnym problemem. Przecież sport, wychowanie fizyczne, rekreacja, rehabilitacja a także wszelka działalność zawodowa wymagają często obydwu rodzajów przejawiania się zdolności motorycznych. Wzbogacone podstawy teoretyczne powinny bowiem znaleźć odzwierciedlenie w zaleceniach metodycznych, zmierzających do zwiększenia efektywności oddziaływania na zdolności motoryczne człowieka.
Dotychczas podejmowano niezbyt liczne próby rozpatrywania współzależności obydwu rodzajów zdolności motorycznych [Pilicz 1986; Szopa 1988, 1992; Starosta 1995a; Mynarski 2000]. Stwierdzono, że istnieją pewne zależności pomiędzy koordynacyjnymi zdolnościami motorycznymi a zdolnościami siłowymi i siłowo-szybkościowymi [Ljach 1990]. Natomiast z badań Wyżnikiewicz-Kopp [1992] wynika, że zależności pomiędzy koordynacyjnymi a kondycyjnymi zdolnościami motorycznymi występowały jedynie sporadycznie. Wydaje się, że u podstaw takiego stanu rzeczy leżą rozmaite rodzaje uwarunkowań koordynacyjnych i kondycyjnych zdolności motorycznych. Pierwsze zależą bowiem od centralnego i obwodowego układu nerwowego. Natomiast drugie w większym stopniu warunkują aspekty morfologiczno-strukturalne i energetyczne [Raczek i wsp. 2002]. Chociaż popularny był i jest nadal pogląd o jedności procesów energetycznych i informacyjnych [Krüger 1989; Vilkner 1989]. Nie przeczy on jednak odrębnemu i analitycznemu rozpatrywaniu a także kształtowaniu tych zdolności.
Wykazano natomiast, że zdolność do przejawiania kompleksowych zdolności motorycznych tj. techniczno-sportowych jest ściśle związana z koordynacyjnymi zdolnościami motorycznymi [Starosta, Hirtz 1989; Raczek 1990]. Jednak pomiędzy zdolnościami koordynacyjnymi a sportowymi zadaniami ruchowymi o strukturze wymagającej wyraźnego przejawienia siły i wytrzymałości występuje niewiele zależności [Raczek 1990]. Podobne związki badano też wcześniej. Przykładem takich poszukiwań mogą być pojęcia „zwinność siłowa” czy „zręczność siłowa” [Regner 1973; Kuzniecov 1975; Ważny 1977].
W badaniach własnych zastosowano zadania motoryczne wymagające globalnego lub lokalnego przejawienia zdolności motorycznych. Dobrano je tak, aby możliwe było szybkie eksponowanie kondycyjnych zdolności motorycznych. Uwzględniono zdolności siłowe i przejawy zdolności szybkościowych w ich trzech podstawowych formach. Analizowano teżzdolność do szybkiego wyzwolenia siły, uznając ją za wypadkową dwóch poprzednich zdolności. Do przejawienia się analizowanych zdolności motorycznych konieczne było rozwinięcie maksymalnych możliwości, niezależnie od charakteru ruchu, który wymagał globalnego lub lokalnego zaangażowania aparatu ruchowego ćwiczącego.
Charakter zadań motorycznych wymagał obciążenia centralnego układu nerwowego i zaangażowania potencjału energetycznego. W tym ostatnim aspekcie uwzględniono wyłącznie mechanizmy beztlenowe. Dlatego w zestawie nie wykorzystano zadań motorycznych wymagających przejawiania wytrzymałości. Skoncentrowano się bowiem nad badaniem efektów krótkotrwałych wysiłków, wymagających szybkiej i adekwatnej reakcji układu nerwowego. Zatem pośrednio analizowano możliwości układu nerwowego i mięśniowego w zakresie maksymalnej aktywności w pojedynczym akcie ruchowym (zadaniu motorycznym) oraz jego ruchliwość w krótkotrwałych zadaniach cyklicznych o maksymalnej lub optymalnej – zadanej częstotliwości. Nie analizowano zmian kondycyjnych i koordynacyjnych zdolności motorycznych pod wpływem narastającego zmęczenia, dlatego że autorzy zajmujący się dotychczas tą problematyką nie wskazywali na powiązania występujące pomiędzy koordynacyjnymi zdolnościami motorycznymi a wytrzymałością [Raczek 1990; Raczek i wsp. 2002]. Chociaż stwierdzono, że wysiłki anaerobowe (30 s – Wingate test) wpływają na poziom zdolności zachowania równowagi, częstotliwość ruchów, poczucie rytmu oraz niektóre przejawy dostosowania motorycznego i sprzężenia ruchów [Waśkiewicz 2002].
Wyboru zadań testowych dokonano na podstawie rozpowszechnionej w ostatnich latach klasyfikacji podstawowych zdolności koordynacyjnych [Raczek i wsp. 2002; Waśkiewicz 2002] (tab. 1).
Poszczególne zdolności koordynacyjne są bardzo trudne do wyodrębnienia i analizy poprzez zastosowanie selektywnych testów. Bardzo często wynik badania zależy równocześnie od dwóch lub więcej zdolności motorycznych. Ten problem wystąpił też w tej pracy. Dlatego niektóre zadania motoryczne powtarzają się jako narzędzia do badania różnych zdolności motorycznych (tab. 1).
W poszczególnych koncepcjach dotyczących zdolności motorycznych stosunkowo niewiele miejsca poświęcono ocenie dokładności wykonania zadań ruchowych. Chociaż, z drugiej strony, większość autorów opracowań dotyczących struktury zdolności motorycznych uznaje zdolność różnicowania ruchu lub zdolność kinestetycznego różnicowania ruchu za jedną z podstawowych koordynacyjnych zdolności motorycznych [Celikovsky i wsp. 1979; Blume 1981; Hirtz 1985; Ljach 1990; Starosta 1993a; Mynarski 2000; Raczek i wsp. 2002]. Stanowi ona zdolność warunkującą odpowiednie wykonanie czynności ruchowych dzięki właściwemu dozowaniu impulsów sterujących, poprzez precyzyjne postrzeganie siłowych, czasowych i przestrzennych parametrów ruchu [Starosta 2001a]. Uważa się też, że zdolność kinestetycznego różnicowania ruchu odgrywa decydującą rolę w powstawaniu „czucia ruchu” [Hotz, Weineck 1983; Starosta 1993a; Hirtz, Starosta 1994; Mynarski 2000].
Tabela 1 Zestawienie zastosowanych w pracy zadań motorycznych na podstawie klasyfikacji podstawowych zdolności koordynacyjnych [Raczek i wsp. 2002; Waśkiewicz 2002]
 |
Pomimo istnienia szeregu opracowań o znaczeniu zdolności różnicowania ruchu, nie dostrzeżono wielu badań dotyczących dokładnego osiągania zadanych parametrów ruchu, a więc uzyskiwania określonej dokładności przejawiania poszczególnych zdolności motorycznych [Starosta 1983, 2001a]. Nie ma też żadnych porównań dokładnego przejawiania zdolności motorycznych w zadaniach ruchowych o globalnym i lokalnym charakterze aktywności. Powstało natomiast wiele prac poświęconych badaniu dostosowania motorycznego, czyli możliwości wdrożenia optymalnego programu działań oraz jego modyfikacji w przypadku dostrzeżenia lub przewidywania zmiany sytuacji [Raczek 1991a]. Zagadnieniami dostosowania motorycznego, które zależy od zdolności różnicowania ruchu, zajmowali się różni autorzy. Jednak prace te miały często charakter teoretyczny [Bernstein 1966; Pöhlmann i wsp. 1979; Czajkowski 1993]. Natomiast rzadko prezentowano wyniki badań eksperymentalnych. Najczęściej prace dotyczyły pojedynczych zdolności lub ogólnych przejawów możliwości sterowania czynnościami motorycznymi [Rutkowska-Kucharska, Bober 1986; Nowacki, Siemieński 1990; Rynkiewicz 1990a, 1990b; Starosta i wsp. 1994; Rynkiewicz, Starosta 2000]. Brakuje natomiast badań nad porównaniem dokładnego przejawiania różnych zdolności motorycznych, szczególnie o globalnym i lokalnym charakterze. Jest to bardzo istotne zagadnienie, ponieważ w sporcie i w życiu codziennym najczęściej niezbędne jest przejawianie ruchów w sposób optymalny, a nie maksymalny. Natomiast większość prac, w których przedmiotem badań były zdolności motoryczne, dotyczyło wysiłku o charakterze maksymalnym.
W badaniach własnych każde z zadań motorycznych stosowano w dwóch wersjach. W pierwszej, głównym celem było osiągnięcie jak największej wartości. W drugiej dążono do sprawdzenia zdolności kinestetycznego różnicowania ruchu jako zewnętrznego przejawu sprawności analizatora ruchowego. Ta ostatnia zdolność jest uznawana za istotną część koordynacji ruchowej, decydującą o wyjątkowej dokładności ruchów [Starosta 1990, 1993a, 2001a]. Natomiast Hirtz [1985] zadania motoryczne wykonywane z maksymalną intensywnością i bez ograniczenia czasowego uznaje za przejawy zdolności do „szybkiej koordynacji” (tab. 2). Umiejętność precyzyjnego wykonania określonej czynności ruchowej jest określana przez Hirtza [1985] jako przejaw „wolnej koordynacji”. Dlatego zadania motoryczne wykorzystywane w pracy podzielono na dwie grupy – zgodne z klasyfikacją Hirtza [1985]. Przyjmując w pierwszej z nich zadania, w których celem było osiągnięcie maksymalnego wyniku. Natomiast w drugiej grupie umieszczono zadania, w których dążono do uzyskania jak największej dokładności, oceniając na tej podstawie zdolność odtwarzania zadanej wielkości ruchu (tab. 2).

Dotychczas w niewielkim zakresie zajmowano się problematyką związków globalnych i lokalnych zdolności motorycznych. Istniejące opracowania dotyczyły głównie ogólnego podziału ruchów na globalne i lokalne [Mĕkota 1986; Ljach 1989, 1990; Starosta 2001a; Mynarski 2000]. Niekiedy określano związki pomiędzy globalnymi a lokalnymi zdolnościami motorycznymi, zwłaszcza w odniesieniu do zdolności koordynacyjnych. Nie podejmowano jednak prób wyjaśnienia rzeczywistych przyczyn istniejących zależności [Ljach 1990].
Brakowało też kompleksowych rozwiązań zagadnień dotyczących relacji występującej pomiędzy kondycyjnymi i koordynacyjnymi zdolnościami motorycznymi. Dotychczasowe dane wskazywały na niejednorodny charakter zależności. Stwierdzono bowiem brak związków pomiędzy koordynacyjnymi zdolnościami motorycznymi a różnymi przejawami wytrzymałości [Havli ek 1972; Mĕkota 1982; Ljach 1990; Raczek i wsp. 1998, 2002]. Nie doszukano się też istotnych współzależności koordynacyjnych zdolności motorycznych ze wskaźnikami siły maksymalnej. Większość współczynników korelacji nie przekraczała 0,30 [Fleischman 1964; Havli ek 1972; Vilkner 1986; Ljach 1990; Turewski 1993]. Ustalono, że w większości przypadków koordynacyjne zdolności motoryczne nie były związane z gibkością [Filipowicz, Turewski 1977; Mĕkota 1982], natomiast były bardziej związane z kondycyjnymi zdolnościami motorycznymi o szybkościowym i siłowo-szybkościowym charakterze [Ljach 1976, 1983, 1990; Turewski 1993; Mĕkota 1982; Vilkner 1986; Raczek i wsp. 1998, 2002].
Istniejący stan wiedzy oraz niebagatelne, teoretyczne i praktyczne, znaczenie tej problematyki było przyczyną podjęcia badań, w których zamierzano określić poziom dwóch podstawowych kondycyjnych zdolności motorycznych, czyli siły i szybkości. Wykonanie wybranych w tym celu zadań motorycznych wymagało przejawienia maksymalnych możliwości. Natomiast przebieg ruchu determinował jego globalne lub lokalne oddziaływanie na ćwiczącego. Zastosowano też zadania motoryczne o złożonej strukturze, wymagające przejawienia wysokiego poziomu różnych zdolności koordynacyjnych.
Zagadnienia związane ze zdolnością różnicowania ruchu są uznawane za podstawowe dla koordynacji ruchowej [Celikovsky i wsp. 1979; Blume 1981; Hotz, Weineck 1983; Hirtz 1985; Ljach 1990; Starosta 1993a, 2001a; Hirtz, Starosta 1994; Mynarski 2000; Raczek i wsp. 2002]. Natomiast nie ma wielu badań dotyczących przejawiania zadanych parametrów ruchu, a więc osiągania określonej dokładności w zakresie poszczególnych zdolności motorycznych [Starosta 1983, 2001a]. Prace najczęściej miały charakter teoretyczny [Bernstein 1966; Pöhlmann i wsp. 1979; Schnabel 1974; Czajkowski 1993]. W zakresie badań eksperymentalnych najczęściej prace dotyczyły pojedynczych zdolności lub ogólnych przejawów możliwości sterowania czynnościami motorycznymi [Rutkowska-Kucharska, Bober 1986; Nowacki, Siemieński 1990; Rynkiewicz 1990a; Starosta i wsp. 1994; Rynkiewicz, Starosta 2000]. Brakowało natomiast badań nad porównaniem dokładnego przejawiania różnych zdolności motorycznych. Ze względu na znaczenie tego zagadnienia podjęto próbę jego kompleksowego rozwiązania. Dlatego zastosowano drugi wariant wykonywania każdego zadania motorycznego. Celem jego realizacji było dokładne, a zatem optymalne wykonanie ruchu. W badaniach w każdym zadaniu motorycznym dążono do uzyskania dokładnie połowy wartości maksymalnej. Przyjęto 50% dlatego, że na tym etapie rozważań nie znano wartości progowych, tzn. nie było możliwe określenie stopnia dokładności różnicowania zadawanych wartości. Nie można było przyjmować wartości zbyt małych ani zbyt bliskich maksymalnym. Istniała bowiem obawa, że badani nie będą potrafili ich zróżnicować. Określano na tej podstawie zdolność kinestetycznego różnicowania ruchu jako zewnętrznego przejawu sprawności analizatora ruchowego. Tę ostatnią zdolność uznaje się za istotną składową koordynacji ruchowej, decydującą o poziomie dokładności ruchów [Blume 1981; Hotz, Weineck 1983; Harre 1985; Starosta 1990, 1993a, 1995a, 2001a; Raczek Mynarski 1992].
Zajmując się rozważaniem nad tymi, w większości nowatorskimi, zagadnieniami,zamierzano osiągnąć następujące cele:
Określenie zależności występujących pomiędzy lokalnymi a globalnymi przejawami kondycyjnych i koordynacyjnych zdolności motorycznych.
Poznanie związków istniejących pomiędzy kondycyjnymi a koordynacyjnymi zdolnościami motorycznymi.
Określenie powiązań pomiędzy optymalnym (dokładnym) i maksymalnym przejawianiem się zdolności motorycznych w kondycyjnych i koordynacyjnych zadaniach o globalnym i lokalnym charakterze oraz stwierdzenie poziomu dokładności wykonywania tych zadań motorycznych.
Ustalenie zakresu symetrii w lokalnym i globalnym przejawianiu się kondycyjnych i koordynacyjnych zdolności motorycznych.
Poszukiwanie dymorfizmu płciowego w zakresie globalnego i lokalnego przejawiania się kondycyjnych i koordynacyjnych zdolności motorycznych.
Dążąc do rozwiązania problemu, przyjęto następujące hipotezy, które zamierzano zweryfikować w zastosowanym procesie badawczym:
Globalne i lokalne zdolności motoryczne są wzajemnie powiązane jedynie w niewielkim stopniu.
Nie ma silnych koordynacyjnych uwarunkowań kondycyjnych zdolności motorycznych, szczególnie o lokalnym charakterze.
Względna wartość błędu w zadaniach wymagających dokładnego przejawiania zdolności motorycznych z zadaną intensywnością uwarunkowana jest ich maksymalnymi wartościami, a możliwości dokładnego wykonywania ruchu zależą od rodzaju zadania motorycznego.
Poziom globalnych i lokalnych przejawów kondycyjnych i koordynacyjnych zdolności motorycznych cechuje znaczny stopień symetryzacji.
W zakresie przejawów zdolności koordynacyjnych kobiety przejawiają poziom podobny do mężczyzn.
Badaniom zrealizowanym w kwietniu i maju 1998 r. poddano studentów III i IV roku studiów stacjonarnych Instytutu Wychowania Fizycznego poznańskiej Akademii Wychowania Fizycznego w Gorzowie Wlkp. Uczestniczyli w nich studenci nie uprawiający wyczynowo sportu od co najmniej dwóch lat. Wszyscy badani byli ludźmi zdrowymi. Każdy z badanych dobrowolnie wyraził zgodę na uczestnictwo w pełnym programie badawczym. Dobierając zadania motoryczne, zwracano uwagę, aby nie były one urazogenne. Nie odbiegały one wymaganiami od typowych ćwiczeń stosowanych w nauczaniu wychowania fizycznego oraz wykonywanych praktycznie na zajęciach z teorii sportu prowadzonych ze studentami.
Przed przystąpieniem do badań wystąpiono o zgodę na ich realizację do Rejonowej Komisji Etyki Badań Naukowych przy Akademii Medycznej im. Karola Marcinkowskiego w Poznaniu. Komisja w dniu 8 kwietnia 1998 r. wyraziła zgodę na wykonanie programu badań.
Badania oparto na jednorodnym materiale złożonym ze studentów AWF z kilku powodów. Pierwszym była możliwość prowadzenia badań na wyselekcjonowanym, jednorodnym materiale, złożonym z osób mających w pełni rozwinięte wszystkie zdolności motoryczne. Kolejnym była możliwość zapewnienia wysokiej motywacji podczas badań, zarówno w zadaniach motorycznych wymagających wysokiej dokładności, jak i w tych, w których konieczne było maksymalne przejawienie zdolności motorycznych. Równie ważne było też to, że studentów III i IV roku AWF można uznać za przedstawicieli populacji, która pozytywnie zakończyła proces edukacji w zakresie wychowania fizycznego. Prawdopodobnie wszystkie analizowane zdolności motoryczne były u badanych w pełni ukształtowane [Hirtz 1978, za Raczek i wsp. 2002]. Natomiast nie rozpoczął się u nich regres zdolności motorycznych spowodowany wiekiem i trybem życia, w którym często brakuje miejsca na kulturę fizyczną, rozumianą jako dbałość o odpowiedni poziom psychomotoryczny. Wyniki uzyskane w badaniach tak dobranej grupy mogą stanowić punkt odniesienia do dalszych rozważań. Osiągnięcie typowego dla badanych studentów poziomu zdolności motorycznych stanowi cel procesu wychowania fizycznego. Z drugiej strony, może być punktem wyjścia w badaniach motoryki osób dorosłych.
Do analizy podstawowych wyników badań wykorzystano dane grupy złożonej ze 176 mężczyzn.
Natomiast rezultaty grupy kobiet, w której znalazły się 84 studentki, rozpatrzono jedynie do oceny dymorfizmu płciowego, przejawianego w zastosowanych zadaniach motorycznych. Średni wiek badanych wynosił odpowiednio 23,2 lata dla mężczyzn oraz 22,7 lat w przypadku kobiet. Charakterystyki masy ciała i jego wysokości przedstawiono w tabeli 3.
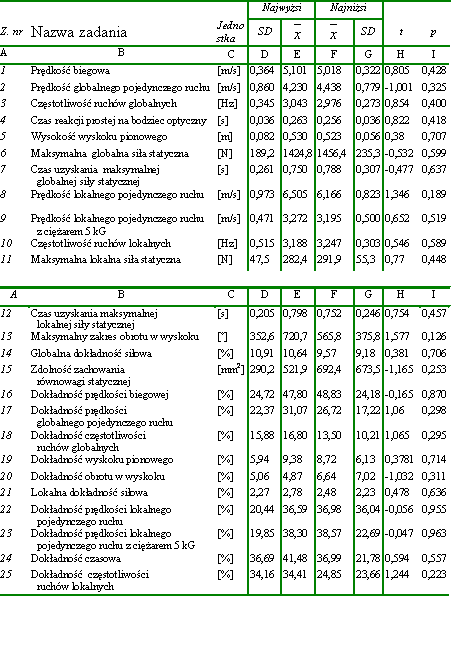
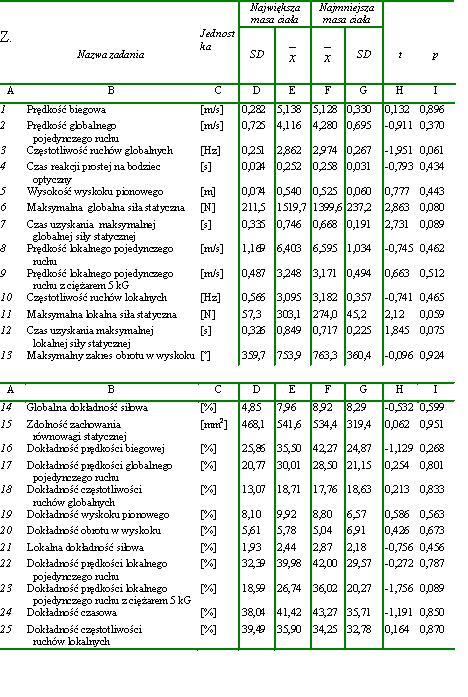
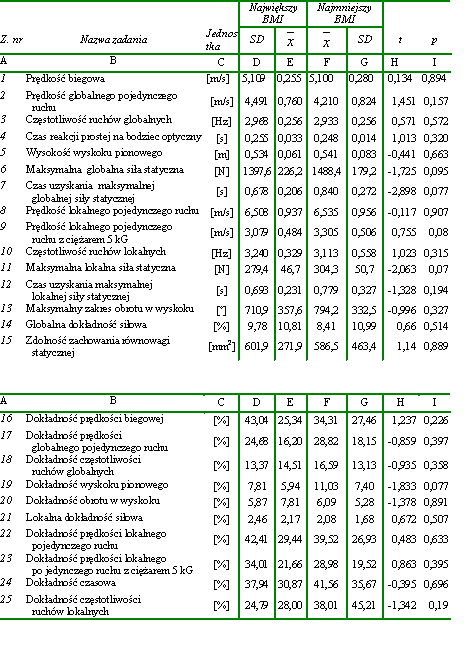
Materiał badawczy wykorzystany w pracy był stosunkowo jednorodny. Współczynniki zmienności dla wysokości i masy ciała wynosiły odpowiednio 3,57% i 6,85% a zatem były stosunkowo niewielkie. W celu potwierdzenia jednorodności dobranego materiału w zakresie badanych zdolności motorycznych, w zależności od wysokości i masy ciała, wyodrębniono spośród wszystkich badanych mężczyzn 30-osobowe grupy. Dwie z nich składały się z osobników najniższych i najwyższych. Natomiast do dwóch kolejnych zakwalifikowano odpowiednio po 30 mężczyzn o najmniejszej i największej masie ciała. Następnie wyliczono istotności różnic pomiędzy uzyskanymi przez nich wynikami w zakresie badanych zdolności motorycznych (tab. 4, tab. 5). Stwierdzono, że żadna z różnic pomiędzy średnimi arytmetycznymi nie była znacząca statystycznie. Dodatkowo obliczono BMI dla wszystkich badanych. Utworzono też dwie grupy 30-osobowe, złożone odpowiednio z osób o najmniejszym i największym BMI (body massindex). Również dokonane w tych grupach porównania średnich arytmetycznych badanych zdolności motorycznych nie upoważniły do wnioskowania o istnieniu różnic znaczących statystycznie (tab. 6). Na tej podstawie uznano materiał za wystarczająco jednorodny dla osiągnięcia celów założonych w pracy.

Tabela 4 Porównanie istotności różnic pomiędzy średnimi arytmetycznymi w obrębie dwóch grup złożonych ze studentów o najniższej (n = 30) i najwyższej (n = 30) wysokości ciała
 |
Spis treści
- 5.1. Pomiar siły i dokładności siłowej
- 5.2. Pomiar prędkości biegowej
- 5.3. Pomiar szybkości reakcji i dokładności czasowej
- 5.4. Pomiar prędkości pojedynczych ruchów
- 5.5. Pomiar wysokości wyskoku pionowego
- 5.6. Pomiar maksymalnego zakresu obrotu w wyskoku
- 5.7. Pomiar częstotliwości ruchów globalnych
- 5.8. Pomiar częstotliwości ruchów lokalnych
- 5.9. Pomiar zdolności zachowania równowagi statycznej
- 5.10. Ocena dokładności przejawiania zdolności motorycznych
- 5.11. Metody statystyczne
- 5.12. Rzetelność stosowanych testowych zadań motorycznych
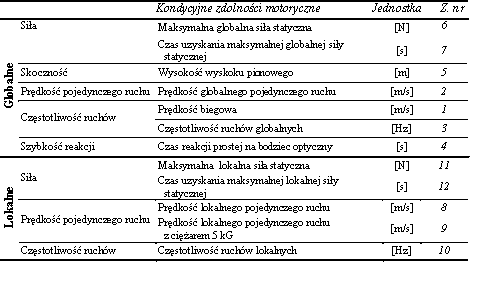
Wyboru zadań testowych do oceny poziomu kondycyjnych zdolności motorycznych dokonano, kierując się ich przydatnością do pomiaru maksymalnej siły statycznej i szybkości. Tę ostatnią zdolność określono w jej różnych elementarnych formach przejawiania się, tj. czasie reakcji, prędkości pojedynczego ruchu i częstotliwości ruchów. Określano globalne i lokalne przejawy wszystkich badanych zdolności. Za pierwsze uznano takie zadania, w których dla prawidłowego wykonania konieczne było zaangażowanie licznych grup mięśniowych i wielu stawów. Natomiast do drugich – lokalnych zakwalifikowano te, w których ruch przebiegał w obrębie jednego stawu [Rutkowska-Kucharska 1982; Rutkowska-Kucharska i wsp.1990] (tab. 7).
Tabela 7 Zestawienie uwzględnionych w badaniach globalnych i lokalnych kondycyjnych zdolności motorycznych
 |
Do oceny koordynacyjnych zdolności motorycznych wykorzystano zadania motoryczne, w których niezbędne było przejawienie szybkiej reakcji i dokładności czasowej lub czasowo-przestrzennej w ruchach pojedynczych oraz cyklicznych (tab. 8). Określano też zdolność zachowania równowagi, stanowiącą jedną z podstawowych zdolności koordynacyjnych o niekwestionowanym znaczeniu praktycznym. Do zestawu zadań wykorzystanych do oceny koordynacji ruchowej włączono także test koordynacji Starosty [1978a]. Dzięki temu uzyskano możliwość porównania wyników testu służącego do kompleksowego mierzenia zdolności koordynacyjnych z rezultatami uzyskanymi w zadaniach motorycznych przeznaczonych do oceny poszczególnych koordynacyjnych zdolności motorycznych [Starosta 1978a] (tab. 8).
Tabela 8 Zestawienie uwzględnionych w badaniach globalnych i lokalnych koordynacyjnych zdolności motorycznych
 |
W stosowanym w badaniach zestawie nie uwzględniono zadań motorycznych wymagających przejawiania wytrzymałości. Skoncentrowano się głównie nad badaniem kondycyjnych i koordynacyjnych zdolności motorycznych w krótkotrwałych wysiłkach, których specyfika wymagała przejawienia szybkiej i adekwatnej reakcji układu nerwowego. A więc, pośrednio analizowano sprawność układu nerwowego i mięśniowego w zakresie maksymalnej aktywności w pojedynczym akcie ruchowym (zadaniu motorycznym) oraz ich ruchliwość w krótkotrwałych zadaniach cyklicznych o maksymalnej lub optymalnej (zadanej) częstotliwości. Celem pracy nie była natomiast analiza zmian poziomu kondycyjnych i koordynacyjnych zdolności motorycznych pod wpływem narastającego zmęczenia.
Ze wszystkich wskaźników uwzględnionych w badaniach wybrano najbardziej reprezentatywne i podzielono je na cztery następujące grupy:
globalne kondycyjne zdolności motoryczne – GKN;
lokalne kondycyjne zdolności motoryczne – LKN;
globalne koordynacyjne zdolności motoryczne – GKO;
lokalne koordynacyjne zdolności motoryczne – LKO.
Przedstawione powyżej skróty przyjęto jako stałe i niezmienne w całej pracy. Podobnie postąpiono w przypadku poszczególnych zadań, gdzie posłużono się numerami, poprzedzonymi skrótem – z. (zadanie) (tab. 9).
Tabela 9 Zestawienie kondycyjnych i koordynacyjnych zdolności motorycznych o globalnym oraz lokalnym charakterze
 |
Podział zdolności motorycznych na kondycyjne i koordynacyjne jest w ujęciu niektórych autorów mało precyzyjny. Dlatego umieszczono w nim jeszcze zdolności szybkościowe jako hybrydowe, uwarunkowane zarówno kondycyjnie, jak i koordynacyjnie [Raczek 1993; Mynarski 2000; Raczek i wsp. 2002].
Dążąc do uproszczenia tego zagadnienia, zaproponowano podział zadań ruchowych na maksymalne i optymalne. Podstawowym kryterium tego podziału stał się zamierzony efekt zadania ruchowego. W maksymalnych przejawach motoryki dążono do uzyskania jak największego, a więc maksymalnego efektu. Natomiast w optymalnych – celem było precyzyjne, czyli najbardziej zbliżone do zaplanowanego zrealizowanie zadania. Miarą poziomu dokładności było odchylenie od wartości zadanej. W tym podziale nie miał żadnego znaczenia stopień złożoności ruchu, czyli podstawowe (często subiektywne) kryterium podziału na kondycyjne i koordynacyjne zadania motoryczne (tab. 10).
Tabela 10 Zestawienie zadań ruchowych zastosowanych do analizy maksymalnych i optymalnych przejawów zdolności motorycznych
 |
Siła mięśniowa stanowi jedną z podstawowych kondycyjnych zdolności motorycznych. Mierzy się ją jako sumę momentów sił mięśniowych lub określa pośrednio na podstawie największej wartości oporu zewnętrznego, pokonanego w określonym zadaniu motorycznym. W badaniach własnych przyjęto drugi sposób pomiaru.
Zastosowano dwa zadania motoryczne. W pierwszym, za pomocą rozciągania dynamometru w leżeniu przodem na poziomej ławeczce, określono globalną siłę statyczną mięśni tułowia i ramion [Wachowski i wsp. 1987]. Badani byli ustabilizowani poprzez unieruchomienie kończyn dolnych na wysokości stawu skokowego. Badani, mając wyprostowane ramiona, trzymali rękoma drążek połączony z dynamometrem. W trakcie pomiaru działali siłą na drążek połączony z dynamometrem.
W drugim zadaniu zmierzono siłę lokalną mięśni zginaczy stawu łokciowego, przy zachowaniu w nim kąta prostego [Starosta i wsp. 1992]. Badani znajdowali się w pozycji siedzącej. W trakcie prowadzenia pomiarów stabilizowano tułów i ramię, dążąc do eliminacji wpływu sąsiednich mięśni. Każdy pomiar wykonywano dwukrotnie, a do obliczeń brano wynik tej próby, w której uzyskano większą siłę. W tym pomiarze badani trzymali uchwyt dynamometru ręką badanej kończyny górnej. Podczas wyzwalania siły, ciągnęli za uchwyt połączony z dynamometrem.
W obydwu zadaniach motorycznych grubość uchwytu była dostosowana do przeciętnych wymiarów dłoni.
Do pomiaru maksymalnej siły statycznej zastosowano dynamometry o zakresie 5000 N – przy pomiarze globalnej siły statycznej i 1000 N – przy pomiarze lokalnej siły statycznej. Tym wartościom odpowiadało napięcie wynoszące 10 000 mV. W pierwszym dynamometrze (do 5000 N) zadziałanie siłą o wartości 1 N powodowało zmianę napięcia o 2 mV. Oznacza to, że dokładność pomiaru wynosiła 0,5 N. Natomiast dla drugiego dynamometru (do 1000 N) odpowiednie wartości wynosiły 10 mV dla 1 N. Umożliwiało to uzyskanie dokładności rzędu 0,1 N. W stosowanym zakresie pomiarów siły urządzenie było liniowe z dokładnością do 2%. Analogowe rezultaty pomiaru siły przetwarzano na zapis cyfrowy poprzez przetwornik analogowo-cyfrowy i w tej postaci zapisywano w pamięci PC. Zapis wartości cyfrowych miał charakter dyskretny, a częstotliwość próbkowania wynosiła 50 herców. Dawało to możliwość określania czasu przejawiania siły z dokładnością do 20 ms, tj. 0,02 s.
Prędkość biegu w przód zmierzono na dystansie 5 m bez rozbiegu, traktując ją jako wskaźnik prędkości startowej [Wachowski i wsp. 1987]. Zachowano jedynie odległość wynoszącą 1,0 m pomiędzy startującym a pierwszą fotodiodą. Postąpiono tak z przyczyn technicznych, zabezpieczając się przed przypadkowym włączeniem pomiaru czasu. Prędkość obliczano na podstawie czasu biegu, który z pomocą aparatury elektronicznej, połączonej z urządzeniami rejestrującymi, mierzono z dokładnością do 0,001 s. Następnie na tej podstawie obliczano prędkość biegu [m/s]. Drogę mierzono taśmą mierniczą (atestowaną) z dokładnością do 0,01 m. Błąd pomiarowy drogi wynosił 0,2 %.
Szybkość reakcji oceniano na podstawie pomiaru czasu reakcji prostej na bodziec optyczny. Założono, że narządem wykonawczym była ręka. Do pomiaru wykorzystano urządzenie sygnalizacyjne sprzężone z elektronicznym miernikiem czasu, funkcjonującym z dokładnością do 0,001 s. Włączniki i wyłączniki pomiaru czasu oraz bodźca świetlnego miały charakterystykę skokową, co zwiększało dokładność pomiaru poprzez eliminację błędu związanego z czasem narastania siły do wartości progowej.
Badanie dokładności czasowej polegało na określaniu czasu wynoszącego 1 s. Badani samodzielnie włączali i wyłączali urządzenie pomiarowe w momencie, w którym uznali, że upłynął zadany odcinek czasu.
W pomiarach obydwu zdolności badani powtarzali zadanie 10 razy, a do analizy przyjmowano sześć wyników pozostałych po odrzuceniu wartości skrajnych. Także w tym przypadku czas rejestrowano miernikiem elektronicznym z dokładnością do 0,001 s.
W tym badaniu zastosowano zestaw urządzeń złożony z dwóch fotodiod sprzężonych z elektronicznym miernikiem czasu, w którym rejestrowano z dokładnością do 0,001 s czasy wykonania ruchów.
Do oceny prędkości pojedynczego ruchu o charakterze lokalnym wykorzystano ruch ramieniem w płaszczyźnie strzałkowej. Podczas realizacji tego zadania motorycznego dolną fotodiodę ustawiano w odległości 1,0 m od podłoża, natomiast górna znajdowała się w położeniu o 0,50 m ponad dolną. Płaszczyzna działania fotodiod była oddalona o 0,60 m od drabinki, przy której ustawiano badanych. Stawali oni tyłem do drabinki, opierając się o nią tułowiem. Następnie, w dowolnie wybranym przez siebie momencie, wykonywali wymach wyprostowanym ramieniem w płaszczyźnie strzałkowej. Podczas badań nie dopuszczano do uginania ramienia w stawie łokciowym. Nie zezwalano też na „odrywanie” tułowia od drabinki. Prędkość pojedynczego ruchu określano na podstawie czasu pokonania odcinka 0,50 m. Ruch ramienia był ruchem obrotowym wokół osi stawu ramiennego. Jednak w badaniach za jego miarę przyjęto wartość pośrednią, jaką była prędkość liniowa rejestrowana w płaszczyźnie działania fotodiod. W każdym zadaniu wykonywano dwa powtórzenia bez dodatkowego obciążenia. Następnie dwa kolejne ruchy realizowano z koniecznością pokonania dodatkowego ciężaru wynoszącego 5,0 kG. Do analizy przyjmowano lepsze rezultaty.
Dla oceny prędkości pojedynczego ruchu o charakterze globalnym wykorzystano prędkość wyprostu (prostowania) tułowia. W tym celu dwie fotodiody ustawiono pionowo jedna nad drugą. Dolna znajdowała się w odległości 0,30 m od podłoża, a górna o 0,50 m wyżej. Badany stał, w skłonie w przód, na wyprostowanych nogach w lekkim rozkroku, bokiem do płaszczyzny, w której działały fotodiody. Podczas pomiaru badany trzymał w rękach drewnianą laskę o długości 2,0 m i średnicy 0,02 m. Jej stosowanie było niezbędne ze względu na konieczność przecinania wiązki światła w celu mierzenia czasu ruchu. Wielkość skłonu uznawano za wystarczającą, jeżeli trzymana laska znajdowała się w odległości 20 cm od podłoża. Następnie w określonym przez siebie momencie wykonywał maksymalnie szybki wyprost tułowia. Zwracano uwagę, aby kończyny dolne były przez cały czas badania wyprostowane w stawach kolanowych. Także w tym pomiarze rejestrowano prędkość liniową, której wartość pośrednio świadczy o wartości rzeczywistej prędkości kątowej ruchu obrotowego tułowia, ramion, szyi i głowy wokół osi obrotu, którą stanowiła oś stawu biodrowego.
Pomiary prędkości pojedynczego ruchu są obarczone pewnym błędem wynikającym z braku możliwości ustabilizowania osi obrotu poruszającego się segmentu ciała (ramienia, tułowia). Wydaje się, że przy stosunkowo niezbyt zróżnicowanym materiale zmiany osi obrotu mogą być zbliżone u poszczególnych badanych. Natomiast bezsporną zaletą tych sposobów pomiaru jest łatwość ich wykonania praktycznie w każdych warunkach, a więc możliwość zastosowania w pomiarach masowych.
Wysokość wyskoku pionowego badano, wykorzystując test skoczności Starosty [Starosta 1978b]. Badani stawali w kole o średnicy 0,80 m, w którego środku umieszczono taśmę mierniczą z naniesioną skalą o dokładności do 0,01 m. Koniec taśmy mocowano poprzez pas do bioder badanego. Pomiar rozpoczynano od określenia wysokości punktu mocowania pasa od podłoża, gdy badany znajdował się w pozycji, stojąc. Następnie w dowolnie wybranym momencie wykonywał, poprzedzony zamachem ramion, wyskok w górę. Jego wysokość określano na podstawie odległości wyciągnięcia taśmy mierniczej. Pomiaru drogi dokonywano z dokładnością do 0,01 m, co mogło powodować 2–3% błąd pomiarowy. Wysokość wyskoku pionowego mierzonego w sposób przyjęty w badaniach własnych wybitnie silnie korelowała z wynikami uzyskanymi na platformie tensometrycznej (r > 0,9) [Radzińska, Starosta 2002].
Wykonywano dwa powtórzenia zadania, a do analizy przyjmowano lepszy wynik. Jeżeli badany po wykonaniu wyskoku nie wylądował stopami w obrębie koła, pomiar powtarzano. Celem takiego postępowania było dokładne określenie wysokości wyskoku pionowego i zmniejszenie błędu spowodowanego poziomym przemieszczeniem się ciała badanego.
Do pomiaru zastosowano test koordynacji Starosty [Starosta 1978a]. Wykorzystano koordynacjomierz Starosty [1978a], który stanowiła drewniana platforma o wielkości 1 m2 z wpisanym kołem o średnicy 0,80 m, na którym znajdowała się skala od 0 do 360 stopni. Badani stawali na środku koła i wykonywali zadanie zgodnie z metodyką podawaną przez Starostę [1978a]. Zadanie rozpoczynano ze ściśle ustalonego położenia stóp (spoczywały one na konturach wyrysowanych na koordynacjomierzu), na których (lewej stopie przy obrocie w lewo, prawej przy obrocie w prawo) zaznacza się kredą linię przechodzącą przez środek pięty oraz miedzy paluchem a drugim palcem. Po wykonaniu maksymalnego obrotu badany musiał zeskoczyć w obrębie czarnego koła, zachowując równowagę, a jego stopa zaznaczała wielkość wykonanego obrotu, którą mierzono, wykorzystując ekierkę i linijkę. Wynik odczytywano z dokładnością do jednego stopnia. Zadanie powtarzano trzykrotnie w prawą i w lewą stronę. Do analizy przyjmowano średnią z wyników trzech pomiarów, oddzielnie dla obrotu w prawo i w lewo. W przypadku niezachowania przez badanego równowagi lub zeskoku poza obręb koła, liczbę powtórzeń zwiększano maksymalnie do pięciu. Badany samodzielnie wybierał kierunek pierwszego wyskoku.
Do wyznaczenia indywidualnych możliwości wykonywania ruchów całego ciała z maksymalną częstotliwością wybrano test Mĕkoty [Mĕkota 1982]. Badani stawali bokiem do listwy o grubości 8 mm i szerokości 20 mm, położonej na podłożu, a następnie wykonywali serię 20 przeskoków obunóż przez nią. Moment rozpoczęcia zadania badani ustalali indywidualnie. Stanowiło to podstawę do rozpoczęcia pomiaru czasu. Natomiast zakończenie jego mierzenia następowało po wykonaniu całego zadania. Częstotliwość określono, dzieląc liczbę przeskoków, tj. 20, przez czas ich wykonania. Określono ją w hercach [cykl/s]. Za jeden cykl uznawano jeden przeskok.
Częstotliwość ruchów lokalnych określono na podstawie czasu wykonania serii 50 powtórzeń – po 25 na każdą stronę – ruchów polegających na jak najszybszym przenoszeniu ręki i dotykaniu krążków umocowanych do blatu, który umieszczono na standardowym biurku. Badani siedzieli na krześle przodem do biurka. W realizacji tego zadania motorycznego przyjęto metodykę typową dla testu EUROFIT [1988, 1989]. Jedynym odstępstwem było zmniejszenie odległości pomiędzy krążkami do 0,40 m. Celem tej modyfikacji było zwiększenie wymagań koordynacyjnych oraz dostosowanie wielkości przestrzeni do realnie występującej, np. klawiatura komputera. Także w tym zadaniu liczbę ruchów dzielono przez czas wykonania zadania i otrzymywano wynik w hercach [cykl/s]. Za jeden cykl uznawano jeden ruch ręki.
Do określenia poziomu zdolności zachowania równowagi zastosowano metodę posturograficzną. Wykorzystano w tym celu system badań „posturograf”, przeznaczony do określania stanu czynnościowego narządu równowagi metodą testów statokinezjometrycznych. Badanie polegało na zachowaniu przez 32 s równowagi w pozycji stojącej na płycie posturograficznej o wymiarach 0,400 m x 0,400 m x 0,055 m. Zespół czujników zamontowany w płycie powoduje rozłożenie siły ciężkości badanego na cztery składowe. Dlatego możliwe jest wyznaczanie chwilowych przemieszczeń rzutu pionowego ogólnego środka ciężkości ciała na platformę [Instrukcja obsługi, 1993]. Badani wykonywali trzy podstawowe zadania motoryczne, polegające na zachowaniu równowagi w pozycji stojąc:
przy oczach otwartych;
przy oczach zamkniętych;
przy oczach otwartych, w warunkach sprzężenia zwrotnego badany obserwował na monitorze aktualną pozycję rzutu ogólnego środka ciężkości (OSC), starając się utrzymać go w centralnym punkcie ekranu.
Przeprowadzona analiza współczynników korelacji pomiędzy poszczególnymi wskaźnikami zdolności zachowania równowagi wykazała, że większość spośród analizowanych wyników była silnie wzajemnie skorelowana. Dlatego do porównań z innymi zdolnościami motorycznymi wybrano pole powierzchni rozwiniętej przez rzut pionowy OSC, traktując go jako wystarczająco reprezentatywny.
Porównano też wyniki typowych dla wszystkich wersji pomiaru zdolności zachowania równowagi. Stwierdzono duże podobieństwo pomiędzy wskaźnikami uzyskiwanymi w różnych sposobach pomiarów. Dlatego analizę ograniczono do wersji podstawowej, tj. pomiaru zdolności zachowania równowagi z oczami otwartymi.
Na podstawie maksymalnych wartości uzyskanych we wszystkich uprzednio wymienionych zadaniach motorycznych obliczano dla każdego badanego, połowy ich wartości. W dalszej kolejności, opierając się na metodzie Schulte – Puni – Starosta [Starosta i wsp. 1992], badani w trzykrotnym powtórzeniu pomiarów uczyli się odtwarzania połowy zadanej wielkości. Następnie w kolejnych dwóch pomiarach dążyli do ich dokładnego odtworzenia. Do analizy przyjmowano wynik tej próby, w której uzyskiwano mniejsze odchylenie od wartości zadanej. Podstawiano go do wzoru i podawano jako bezwzględną wartość błędu wyrażoną w procentach.
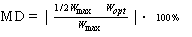 (1)
(1)
gdzie:
MD – miara dokładności, wyrażona w % od wartości zadanej, tj. 50% wartości maksymalnej;
Wmax – wynik maksymalny w danym powtórzeniu zadania motorycznego;
Wopt – wynik uzyskany w zadaniu motorycznym wymagającym przejawienia dokładności, tj. uzyskania połowy wartości maksymalnej.
Dzięki takiemu postępowaniu, którego efektem była miara dokładności wyrażona w procentach, uzyskano możliwość porównywania odchyleń od wartości zadanej dla różnych zdolności motorycznych, mierzonych w rozmaitych jednostkach. Wartości odchyleń od niej uznawano za miarę poszczególnych rodzajów dokładności, np. dla maksymalnej siły globalnej i lokalnej były to wskaźniki globalnej i lokalnej dokładności siłowej.
Przed przystąpieniem do opisania metod statystycznych, zastosowanych w analizie wyników badań własnych, dokonano charakterystyki dwóch współczynników, których obliczenie i analiza były niezbędne do osiągnięcia niektórych celów badań.
Możliwości przejawiania zdolności motorycznych u kobiet i mężczyzn porównano, posługując się wzorem, który umożliwiał określenie różnic pomiędzy identycznymi czynnościami ruchowymi wykonywanymi przez studentki i studentów, z uwzględnieniem wartości średnich arytmetycznych i średnich odchyleń standardowych [Szopa i wsp. 1985].
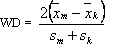 (2)
(2)
gdzie:
WD – wskaźnik dymorfizmu;
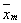 – średnia arytmetyczna zadania wykonywanego przez
mężczyzn;
– średnia arytmetyczna zadania wykonywanego przez
mężczyzn;
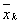 – średnia arytmetyczna zadania wykonywanego przez
kobiety;
– średnia arytmetyczna zadania wykonywanego przez
kobiety;
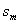 – odchylenie standardowe zadania wykonywanego
przez mężczyzn;
– odchylenie standardowe zadania wykonywanego
przez mężczyzn;
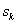 – odchylenie standardowe zadania wykonywanego
przez kobiety.
– odchylenie standardowe zadania wykonywanego
przez kobiety.
Zbliżony sposób postępowania przyjęto dla określenia wskaźnika symetryzacji, który jest miarą podobieństwa przejawów zdolności motorycznych w czynnościach wykonywanych prawą i lewą kończyną lub w prawą i lewą stronę. Zastosowano wzór skonstruowany na podobieństwo zaproponowanego przez Szopę, Mleczko i Cemplę [1985] do oceny dymorfizmu płciowego:
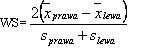 (3)
(3)
gdzie:

Uzyskane wyniki opracowano statystycznie, wykorzystując program „STATISTICA 6,0 PL”. Wyboru metod do obliczania danych dokonano, mając na uwadze konieczność osiągnięcia podstawowych celów, a zatem zbadania zależności pomiędzy zdolnościami motorycznymi, wyszukania podobieństw między nimi i ich wzajemnych uwarunkowań oraz określenia różnic w przejawach motoryczności w symetrycznych zadaniach ruchowych oraz pomiędzy grupami kobiet i mężczyzn.
Stwierdzono, że wszystkie wartości przyjęte do tej analizy cechowały się rozkładem normalnym. Wyliczono średnie arytmetyczne oraz istotności różnic między nimi. Obliczono też macierze korelacji pomiędzy analizowanymi zdolnościami motorycznymi [Greń 1976; Furdal 1989].
Dążąc do określenia wzajemnych uwarunkowań poszczególnych zdolności motorycznych, obliczano regresje wielokrotne, w których kolejne zadania motoryczne traktowano jako zmienną zależną (objaśnianą), a pozostałe były zmiennymi niezależnymi (objaśniającymi). Aby uzyskać informacje o rzeczywistym charakterze i sile wzajemnych uwarunkowań zdolności motorycznych, dokonano w każdym modelu redukcji równań regresji wielokrotnej, posługując się metodą regresji krokowej wstecznej, którą kontynuowano aż do pełnej redukcji, czyli do uzyskania maksymalnie zredukowanego równania. Założono w tych działaniach minimalne wartości progowe do wprowadzenia i do usunięcia, tak aby w równaniach można było uwzględnić jak najwięcej zmiennych objaśniających.
Dążąc do określenia podobieństw pomiędzy analizowanymi zmiennymi, dokonano ich grupowania. Wykorzystano w tym celu metodę aglomeracji cech. Podczas formowania skupień zastosowano najbardziej bezpośredni sposób określania odległości między obiektami w przestrzeni wielowymiarowej, polegający na obliczaniu odległości euklidesowej, tj. odległości geometrycznej w przestrzeni wielowymiarowej. Posłużono się w tym celu następującym wzorem:
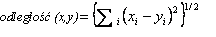 (4)
(4)
Następnie odległość euklidesową podniesiono do kwadratu, aby przypisać większą wagę bardziej oddalonym zmiennym. Obliczono ją następująco:
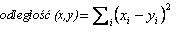 (5)
(5)
Aglomeracji cech dokonano, stosując metodę Warda, która umożliwia wykorzystanie analizy wariancji do oszacowania odległości między skupieniami. Dąży się w niej do minimalizacji sumy kwadratów dowolnych dwóch skupień, które mogą zostać uformowane na każdym etapie [Ward 1963]. Jest ona uznawana za bardzo efektywną, chociaż zmierza do tworzenia skupień o małej wielkości
W efekcie uzyskano poziomy hierarchiczny wykres drzewkowy. W jego początkowej formie każda zmienna stanowi swoją własną klasę. Metoda polega na sukcesywnym, dokonywanym małymi krokami, „osłabianiu” kryterium oceniającego wyjątkowość zmiennych lub ich brak. Innymi słowy, działanie polega na ciągłym obniżaniu progu stanowiącego o decyzji przypisania dwóch lub więcej zmiennych do tego samego skupienia. Tym sposobem wiąże się ze sobą coraz to więcej zmiennych i agreguje je w coraz to większe skupienia elementów coraz bardziej różniących się od siebie. W końcu, na ostatnim etapie, wszystkie obiekty zostają ze sobą połączone. Na wykresach tych na osi poziomej odłożone są odległości aglomeracyjne. Zatem przy każdym węźle na wykresie (gdzie uformowało się nowe skupienie) możemy odczytać odległość, w której odpowiednie elementy zostały powiązane ze sobą, tworząc nowe pojedyncze skupienie. Jeśli dane mają wyrazistą „strukturę” w tym sensie, że istnieją skupienia podobnych do siebie obiektów, to często struktura ta znajdzie odbicie na hierarchicznym drzewie w postaci oddzielnych gałęzi. Pomyślna analiza za pomocą metody łączenia daje możliwość wykrywania skupień (gałęzi) i ich interpretacji.
Jednym z podstawowych kryteriów, wykorzystywanych do oceny przydatności zadań motorycznych do oceny zdolności motorycznych, jest ich rzetelność. Mĕkota [1986] definiuje ją jako trafność testu wobec samego siebie. Najczęściej mierzy się ją poprzez obliczanie współczynnika korelacji pomiędzy wynikami dwóch, powtórzonych pomiarów [Osiński 2000]. Według Zaciorskiego [1979] w diagnostyce zdolności motorycznych powinno się uwzględniać następujące kryteria współczynników rzetelności:
0,95 – 0,99 – rzetelność doskonała;
0,90 – 0,94 – rzetelność dobra;
0,80 – 0,90 – rzetelność dopuszczalna;
0,70 – 0,79 – rzetelność bardzo słaba;
0,60 – 0,69 – rzetelność wątpliwa dla ocen indywidualnych.
W badaniach zdolności motorycznych zazwyczaj przyjmuje się za wystarczającą wartość współczynnika korelacji wynoszącą 0,70 [Szczotka 1969; Zaciorski 1979; Osiński 1993]. Jej uzyskanie jest szczególnie problematyczne w pomiarach koordynacyjnych zdolności motorycznych, które są bardziej złożone niż ich kondycyjne przejawy [Mynarski 2000]. Testy koordynacyjne najczęściej polegają na wykonywaniu krótkotrwałych zadań. Dlatego nie ma w nich możliwości kompensowania błędu pomiarowego przez inne czynniki [Neumeier 1983; Blume, Zimmermann 1987]. Lienert [1969, za Mynarskim 2000] twierdził, że w testach psychomotorycznych współczynnik rzetelności 0,50 jest wystarczający do diagnozy grupowej. Wiszniakow [1993] uważał, że próby, w których konieczne jest uzyskanie znacznej dokładności, mają współczynniki rzetelności mieszczące się w przedziale od 0,3 do 0,7. Miary rzetelności zadań motorycznych zastosowanych w badaniach własnych przedstawiono w tabeli 11. Uzyskane wyniki mieściły się w ogólnie przyjętych przedziałach wartości.

Część zastosowanych w pracy zadań testowych charakteryzowała się stosunkowo niskimi wartościami rzetelności, które jednak nie odbiegały od uznawanych za dopuszczalne w tego typu badaniach. Dlatego tych prób nie odrzucono. Jednak uzyskane wyniki należy traktować stosunkowo ostrożnie.
Ryzyko mało rzetelnej oceny zdolności motorycznych można zmniejszyć, stosując odpowiednią procedurę podczas badań. Według Zaciorskiego [1979] zwiększeniu rzetelności może sprzyjać:
bardziej rygorystyczna standaryzacja;
zwiększenie liczby prób;
zwiększenie liczby oceniających oraz poprawa ich umiejętności w zakresie oceny zjawisk;
wprowadzenie testów ekwiwalentnych;
zwiększenie motywacji badanych.
W badaniach własnych uwzględniono przedstawione powyżej wymagania. Wszystkie pomiary wykonywano w warunkach wystandaryzowanych. Zadania motoryczne miały charakter ilościowy, a nie jakościowy. W większości zadań testowych wykorzystano elektroniczną aparaturę kontrolno-pomiarową. Liczbę powtórzeń poszczególnych zadań motorycznych dostosowano do spodziewanej rzetelności. Wykonywano od 2 do 5 powtórzeń zadania, a do analizy wybierano lepszy wynik. W pomiarach czasu reakcji i dokładności czasowej zastosowano 10 powtórzeń. Dążąc do eliminacji przypadkowych rezultatów, odrzucano w nich po dwie wartości skrajne – dwie minimalne i dwie maksymalne.
Dla zapewnienia wysokiej motywacji przy wykonywaniu zadań motorycznych, zapewniono znaczące nagrody o charakterze prestiżowym dla dziesięciu badanych osiągających najlepsze (maksymalne) wyniki i dziesięciu najbardziej dokładnych. Ich listę rangową ustalono na podstawie sumarycznych wyników wykonania wszystkich zadań. W jednym zestawieniu znaleźli się osobnicy uszeregowani ze względu na maksymalne osiągnięcia. Natomiast w drugim sklasyfikowano badanych według największej dokładności odtworzenia zadanych wartości. Identyczne kryteria zastosowano dla kobiet i mężczyzn.
Kolejnym zagadnieniem, które pominięto w pracy, jest badanie trafności zadań testowych. Zastosowane w pracy zadania testowe były badane pod względem trafności do oceny kondycyjnych zdolności motorycznych [Wachowski, Strzelczyk 1999]. Uzyskane przez wymienionych autorów wyniki dowodziły bardzo dużej trafności tych testów. O wiele bardziej złożonym problemem jest badanie trafności zadań testowych wykorzystanych do oceny koordynacyjnych zdolności motorycznych. Pierwszym, zasadniczym problemem jest brak powszechnej zgodności co do ilości i rodzaju koordynacyjnych zdolności motorycznych. Drugim, wynikającym z pierwszego, utrudnieniem jest brak niekwestionowanych sposobów oceny poszczególnych koordynacyjnych zdolności motorycznych. Brakuje zatem testów, do których można odnieść trafność zadań zastosowanych w pracy. Brak kompletnej analizy trafności zastosowanych w pracy zadań testowych zmusza zatem do bardziej ostrożnego wnioskowania.
Spis treści
- 6.1. Zależności pomiędzy kondycyjnymi zdolnościami motorycznymi
- 6.2. Zależności pomiędzy koordynacyjnymi zdolnościami motorycznymi
- 6.3. Zależności pomiędzy kondycyjnymi a koordynacyjnymi zdolnościami motorycznymi
- 6.4. Lokalne a globalne zdolności motoryczne
- 6.5. Kondycyjne a koordynacyjne zdolności motoryczne
- 6.6. Charakterystyka podobieństw pomiędzy badanymi zdolnościami motorycznymi
- 6.7. Maksymalne i optymalne przejawy zdolności motorycznych o globalnym i lokalnym charakterze
- 6.8. Czas narastania siły w zadaniach o maksymalnym i optymalnym charakterze jako wyraz specyficznych wymogów związanych z dokładnością siłową
- 6.9. Zdolność do dokładnego odtwarzania zadanej wartości zdolności motorycznych a rodzaj ruchu
- 6.10. Symetryczność globalnych i lokalnych przejawów kondycyjnych oraz koordynacyjnych zdolności motorycznych
- 6.11. Zróżnicowanie płciowe globalnych i lokalnych przejawów kondycyjnych i koordynacyjnych zdolności motorycznych
W pierwszej kolejności dążono do określenia wzajemnych relacji w obrębie uwarunkowań globalnych kondycyjnych zdolności motorycznych. Uzyskane wyniki w postaci podstawowych charakterystyk statystycznych przedstawiono w tabeli 12. Następnie poddano je dalszej analizie, której celem było ustalenie statystycznie znaczących współczynników korelacji pomiędzy wynikami wykonania poszczególnych zadań motorycznych (tab. 13).

W obrębie zadań motorycznych wykorzystanych do określenia globalnych kondycyjnych zdolności motorycznych stwierdzono jedynie nieliczne powiązania. Prędkość wyprostu, która warunkowała zdolność do szybkiego wykonania pojedynczego ruchu (z. nr 2), zależała od częstotliwości przeskoków (z. nr 3) oraz od minimalnego czasu uzyskania maksymalnej globalnej siły (z. nr 7). Wydaje się zatem, że osobnicy cechujący się zdolnościami szybkiego wykonania pojedynczych ruchów dysponowali także zdolnością do realizacji ruchów o znacznej częstotliwości oraz umiejętnością szybkiego osiągania maksymalnej siły (tab. 13).
Tabela 13 Statystycznie istotne współczynniki korelacji w obrębie globalnych kondycyjnych zdolności motorycznych (n = 176)
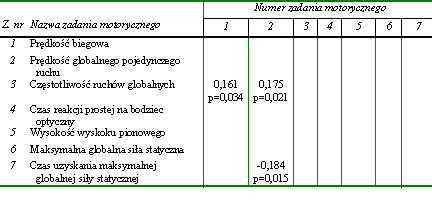 |
Wszystkie współczynniki korelacji pomimo niewielkich wartości, mieszczących się w przedziale od 0,161 do -0,184, były wysoce znaczące statystycznie (tab. 13). Jednak ich niewielka liczba oraz niskie wartości dowodziły niezbyt silnych zależności pomiędzy poszczególnymi globalnymi kondycyjnymi zdolnościami motorycznymi. Wydaje się, że większość zadań motorycznych wykorzystanych w badaniach do oceny globalnych kondycyjnych zdolności motorycznych miała względnie niezależny charakter.
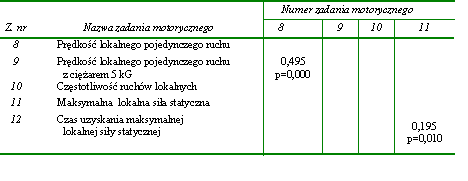
Statystycznie istotne związki pomiędzy rezultatami zadań motorycznych zastosowanych do oceny lokalnych przejawów kondycyjnych zdolności motorycznych stwierdzono jedynie w dwóch przypadkach. Pierwszy dotyczył prędkości lokalnego pojedynczego ruchu wykonywanego bez obciążenia zewnętrznego (z. nr 8) i z obciążeniem ciężarem 5 kG (z. nr 9) (tab. 14). Drugi stwierdzono pomiędzy wielkością maksymalnej lokalnej siły statycznej (z. nr 11) a minimalnym czasem niezbędnym do jej przejawienia (z. nr 12) (tab. 14). Współczynniki korelacji wynosiły odpowiednio 0,495 i 0,195 (tab. 14).
Na podstawie przedstawionych wyników można wnioskować o istnieniu bardzo niewielu powiązań w obrębie przejawów globalnych i lokalnych kondycyjnych zdolności motorycznych. Można zatem stwierdzić, że zadania motoryczne zastosowane do oceny lokalnych kondycyjnych zdolności motorycznych były względnie niezależne. Natomiast pojedyncze istotne statystycznie związki nie upoważniają do wnioskowania o silnym wzajemnym powiązaniu kondycyjnych zdolności motorycznych tak o globalnym, jak i lokalnym charakterze przejawiania się. Tym bardziej że odnotowane istotne współczynniki korelacji nie były wysokie.
W dalszej kolejności poszukiwano związków pomiędzy wynikami zadań motorycznych wykorzystanych do oceny globalnych i lokalnych kondycyjnych zdolności motorycznych. Także w tym przypadku zależności nie było zbyt wiele. Najsilniejsze współczynniki korelacji stwierdzono pomiędzy maksymalną lokalną siłą statyczną (z. nr 11) i maksymalną globalną siłą statyczną (z. nr 6) (r = 0,516) a także pomiędzy czasami niezbędnymi do uzyskania obydwu wartości siły (z. nr 12 z z. nr 7) (r = 0,434) (tab. 15). Można na tej podstawie przypuszczać, że siła statyczna była zdolnością niespecyficzną, przejawiającą się niezależnie od rodzaju zadania ruchowego. Wystąpiła też istotna statystycznie zależność pomiędzy częstotliwością ruchów o charakterze globalnym (z. nr 3) i lokalnym (z. nr 10). Prawdopodobnie także za niespecyficzną można uznać zdolność szybkiego przechodzenia układu nerwowego ze stanu pobudzenia do hamowania i odwrotnie. Jest ona podstawowym czynnikiem determinującym uzyskiwanie dużej częstotliwości ruchów. Jest ona także niezależna od rodzaju ruchu. Podobnie jak w poprzedniej grupie zdolności motorycznych prędkość wykonania pojedynczego ruchu ramieniem (z. nr 2) zależała od umiejętności szybkiego przejawienia maksymalnej lokalnej siły statycznej (z. nr 12) (tab. 15).
Istotne współczynniki korelacji zaobserwowano też w przypadku prędkości pojedynczych ruchów o globalnym (z. nr 2) i lokalnego przejawiania się aktywności mięśniowej (z. nr 8) a także pomiędzy tą zdolnością (z. nr 8) a prędkością biegową (z. nr 1) i częstotliwością przeskoków (z. nr 3) (tab. 15).
Na podstawie wyników dokonanej analizy współczynników korelacji można sądzić, że możliwości przejawiania kondycyjnych zdolności motorycznych, określone za pomocą zastosowanych zadań motorycznych, cechowało pewne podobieństwo w zakresie ich globalnego i lokalnego przejawiania. Chociaż każdy rodzaj kondycyjnych zdolności motorycznych wydawał się być względnie niezależny. Należy podkreślić, że niezbyt wysokie (chociaż znaczące statystycznie) współczynniki korelacji skłaniają do znacznej ostrożności w określaniu istniejących powiązań pomiędzy poszczególnymi zdolnościami motorycznymi (tab. 15).
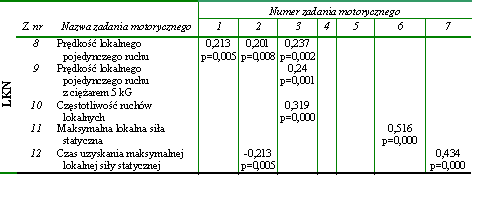
Podobnie jak w poprzedniej grupie zadań motorycznych, obliczono średnie arytmetyczne z uzupełnieniami charakteryzujące globalne i lokalne koordynacyjne zdolności motoryczne (tab. 16). Następnie określono współczynniki korelacji liniowej pomiędzy wynikami wykonania poszczególnych zadań motorycznych (tab. 17, tab. 18).

Globalne koordynacyjne zdolności motoryczne określone na podstawie zastosowanych zadań motorycznych nie były znacząco wzajemnie związane. Stwierdzono jedynie pięć przypadków niskich, ale statystycznie istotnych współczynników korelacji (tab. 17). Wynik maksymalnego obrotu w wyskoku mierzony testem Starosty (z. nr 13) był znacząco skorelowany z dokładnością prędkości biegowej (z. nr 16) i polem powierzchni rozwiniętej przez rzut pionowy OSC w pomiarze zachowania równowagi w pozycji stojąc na posturografie (z. nr 15). Zależności te miały charakter ujemny, co oznaczało, że osobnicy o wyższym poziomie zdolności zachowania równowagi oraz większej dokładności ruchów w odtwarzaniu zadań lokomocyjnych osiągali lepsze wartości obrotu w wyskoku. Zadania oznaczone numerami 15 i 16, tj. zdolność zachowania równowagi i dokładność w cyklicznych ruchach lokomocyjnych, były wzajemnie skorelowane (tab. 17).
Tabela 17 Statystycznie istotne współczynniki korelacji w obrębie globalnych koordynacyjnych zdolności motorycznych (n = 176)
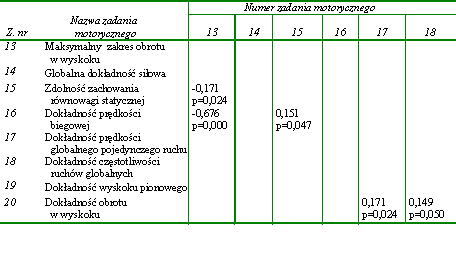 |
Interesujące, że wystąpiły niewielkie, chociaż znaczące powiązania pomiędzy precyzyjnym wykonaniem pojedynczego ruchu tułowia (z. nr 17) a umiejętnościami odtworzenia zadanej wartości obrotu w wyskoku (z. nr 20) (tab. 17). Współczynnik był istotny na poziomie wynoszącym p = 0,05.
Na podstawie analizy współczynników korelacji pomiędzy rezultatami zastosowanych zadań motorycznych lokalne koordynacyjne zdolności motoryczne w zasadzie można uznać za wzajemnie niezależne. Jedynym wyjątkiem było stwierdzenie wybitnie silnego związku pomiędzy dokładnością czasową (z. nr 24) a umiejętnością odtworzenia połowy częstotliwości ruchów kończyną górną (z. nr 25). Wartość współczynnika korelacji wynosiła tu aż 0,714 (tab. 18).

Podczas analizy zależności pomiędzy globalnymi a lokalnymi koordynacyjnymi zdolnościami motorycznymi określono jedynie dwa przypadki statystycznie istotnych powiązań pomiędzy zdolnościami z obydwu grup (tab. 19). I tak lokalna dokładność siłowa (z. nr 21) była skorelowana z możliwościami dokładnego odtworzenia połowy wysokości maksymalnego pionowego wyskoku (z. nr 19). Natomiast drugi, znaczący współczynnik korelacji, stwierdzono pomiędzy dokładnym odtworzeniem zadanej wartości częstotliwości ruchów o globalnym i lokalnym charakterze (z. nr 18 oraz z. nr 25).
Na podstawie przeprowadzonej analizy współczynników korelacji można wnioskować o większej niezależności koordynacyjnych zdolności motorycznych, szczególnie o lokalnym charakterze. Prawdopodobnie mechanizmy decydujące o umiejętnościach precyzyjnego wykonywania rozmaitych lokalnych zadań ruchowych były bardzo specyficzne i różnorodne.
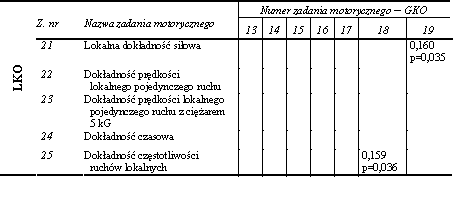
Poszukując powiązań pomiędzy wynikami zadań motorycznych zastosowanych do analizy kondycyjnych i koordynacyjnych zdolności motorycznych, nie stwierdzono zbyt wielu znaczących statystycznie współczynników korelacji (tab. 20). Trzy zdolności koordynacyjne, tj.:
dokładność prędkości biegowej – z. nr 16;
dokładność obrotu w wyskoku – z. nr 20;
dokładność prędkości lokalnego pojedynczego ruchu z ciężarem 5 kG – z. nr 23;
oraz dwie zdolności kondycyjne, którymi były:
wysokość wyskoku – z. nr 5;
minimalny czas uzyskania maksymalnej lokalnej siły statycznej – z. nr 12; nie były w żaden, statystycznie istotny sposób, powiązane z innymi zdolnościami (tab. 20).
Możliwość wykonania znacznego obrotu w wyskoku (z. nr 13) zależała dodatnio od prędkości lokalnego pojedynczego ruchu (z. nr 8, z. nr 9) oraz ujemnie od czasu reakcji prostej na bodziec optyczny (z. nr 4). Przypuszczalnie o poziomie maksymalnego obrotu w wyskoku decydowała zarówno zdolność do szybkiej reakcji na nagle zmieniające się warunki, jak też umiejętność precyzyjnego sterowania ułożeniem części ciała w przestrzeni (tab. 20).
Zarówno globalna jak i lokalna dokładność siłowa (z. nr 14; z. nr 21) zależały od maksymalnej globalnej siły statycznej (z. nr 6). Ujemny współczynnik korelacji sugerował, że osobnicy silniejsi byli też bardziej precyzyjni – popełniający mniejsze błędy. Maksymalna globalna siła statyczna (z. nr 6) była także ujemnie skorelowana z dokładnością wyskoku pionowego (z. nr 19), który można uznać za przejaw dokładności w zakresie przejawiania skoczności (tab. 20).
Zdolność zachowania równowagi (z. nr 15), określana na podstawie wielkości pola powierzchni rozwiniętej przez rzut pionowy OSC, była ujemnie związana z czasem przejawiania maksymalnej globalnej siły statycznej (z. nr 7) (tab. 20).
Interesujące zjawisko stwierdzono podczas analizy związków, które wystąpiły pomiędzy częstotliwością ruchów o globalnym i lokalnym charakterze (z. nr 3, z. nr 10) oraz umiejętnością odtworzenia zadanej częstotliwości (czyli dokładnością częstotliwości) tych ruchów (z. nr 18; z. nr 25). W zadaniu o charakterze globalnym (przeskoki obunóż – test Mĕkoty) zależność ta była dodatnia (z. nr 3, z. nr 18), co dowodziło, że osobnicy przejawiający większą częstotliwość byli mniej precyzyjni w ruchach globalnych. Natomiast w zadaniach o lokalnym charakterze (ruchy kończyną górną – EUROFIT) stwierdzono odwrotną tendencję (z. nr 10, z. nr 25) (tab. 20).
Niejednorodny charakter związków w obrębie lokalnych i globalnych zdolności motorycznych stwierdzono także w zakresie prędkości pojedynczych ruchów i dokładności przejawienia ich zadanej wartości. Statystycznie znaczącą zależność zaobserwowano jedynie w zakresie zadania o charakterze globalnym (z. nr 2 i z. nr 22) (tab. 20). Interesujące, że prędkość wyprostu tułowia (z. nr 3), traktowana jako miara zdolności do wykonania pojedynczego ruchu o globalnym charakterze, była dodatnio związana z dokładnością odtworzenia zadanej częstotliwości ruchów globalnych (z. nr 18). Sugerowało to mniejszą dokładność wykonania ruchów z zadaną częstotliwością u osobników o znacznych możliwościach szybkiego wykonywania globalnego pojedynczego ruchu. Natomiast dokładność czasowa (z. nr 24) korelowała ujemnie z częstotliwością ruchów o lokalnym charakterze (z. nr 10). W tym ostatnim przypadku badani o większych zdolnościach do wykonywania ruchów lokalnych z dużą częstotliwością przejawiali też większą dokładność podczas wykonywania serii ruchów kończyną górną z zadaną częstotliwością (z. nr 10 i z. nr 25) (tab. 20).
Podczas analizy uzyskanych wyników należy pamiętać, że większość współczynników korelacji uznanych za istotne nie miała zbyt wysokich wartości. Dlatego uzyskane wyniki należy traktować z pewną dozą ostrożności.
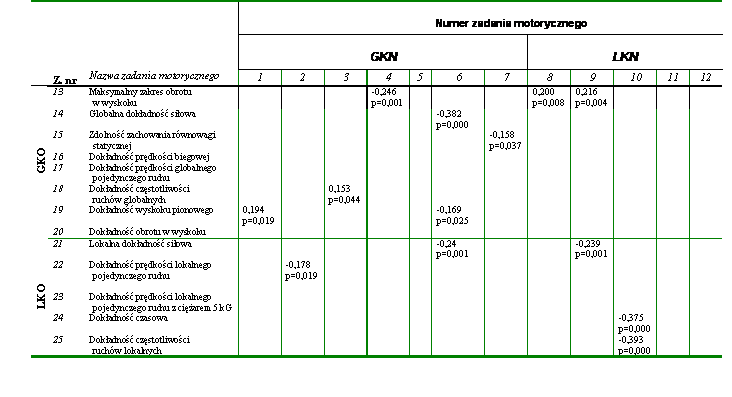
Analiza współczynników korelacji nie upoważniła do wnioskowania o istnieniu silnych i pewnych zależności pomiędzy globalnymi a lokalnymi przejawami kondycyjnych i koordynacyjnych zdolności motorycznych. Nie było też podstaw do określenia siły tych związków. Dane uzyskane po rozpatrzeniu współczynników korelacji nie były jednoznaczne i silne. Dlatego posłużono się metodą analizy regresji wielokrotnej z wyborem optymalnego podzbioru zmiennych niezależnych. Przypuszczano, że bardziej precyzyjna metoda umożliwi określenie rzeczywistej siły i charakteru związków w obrębie mierzonych zdolności motorycznych oraz ułatwi eliminację przypadkowych współczynników korelacji, świadczących o pozornych a nie rzeczywistych zależnościach pomiędzy nimi.
]Jedynie trzy spośród analizowanych globalnych kondycyjnych zdolności motorycznych zależały od lokalnych przejawów kondycyjnych zdolności motorycznych. W tych przypadkach redukcja modeli regresji wielokrotnej zakończyła się uzyskaniem równania znaczącego statystycznie. Najsilniejszy związek wystąpił pomiędzy globalną maksymalną siłą statyczną (z. nr 6) a jej lokalnym odpowiednikiem (z. nr 11). Współczynnik korelacji wielokrotnej wynosił 0,516 (tab. 21).
Podobną, chociaż nieco słabszą zależność określono w równaniu, gdzie zmienną zależną był czas osiągania maksymalnej globalnej siły statycznej (z. nr 7). Spośród zmiennych niezależnych po redukcji pozostał czas uzyskania maksymalnej lokalnej siły statycznej (z. nr 12), a współczynnik korelacji takiego równania wynosił 0,421. Ostatnią globalną kondycyjną zdolnością motoryczną, która zależała od lokalnych przejawów motoryczności, była częstotliwość ruchów o charakterze globalnym (z. nr 3), określona zdolnością do szybkiego wykonywania serii przeskoków. Jej wartość była uwarunkowana częstotliwością ruchów o lokalnym charakterze (z. nr 10) oraz prędkością pojedynczego ruchu kończyną górną (pojedynczy ruch lokalny) (z. nr 8). Współczynnik korelacji wielokrotnej maksymalnie zredukowanego modelu regresji wynosił 0,400. Natomiast współczynniki BETA charakteryzujące jego składowe osiągnęły wartości wynoszące odpowiednio 0,323 i 0,241 (tab. 21). Wszystkie powiązania miały charakter dodatni, co dowodziło istnienia wprost proporcjonalnej zależności pomiędzy analizowanymi zdolnościami motorycznymi. Związki uzyskane podczas analizy regresji określono też w analizie współczynników korelacji pomiędzy globalnymi a lokalnymi kondycyjnymi zdolnościami motorycznymi.

Redukcja równań regresji wielokrotnej, gdzie zmiennymi zależnymi były globalne koordynacyjne zdolności motoryczne a zmiennymi niezależnymi ich lokalne przejawy, doprowadziła do wnioskowania o braku związków pomiędzy nimi. Żadne z równań regresji wielokrotnej nie było znaczące statystycznie. Ich redukcja nie umożliwiła wskazania jakichkolwiek powiązań pomiędzy globalnymi a lokalnymi koordynacyjnymi zdolnościami motorycznymi. Stanowiło to potwierdzenie wyników analizy współczynników korelacji, w której uzyskano jedynie dwa i to bardzo niskie, istotne współczynniki korelacji.
Dążąc do określenia powiązań pomiędzy kondycyjnymi a koordynacyjnymi zdolnościami motorycznymi, przeprowadzono analizę regresji wielokrotnej, gdzie zmiennymi zależnymi były zdolności kondycyjne a zmiennymi niezależnymi zdolności koordynacyjne. Aby wyznaczyć zdolności kondycyjne i koordynacyjne najsilniej wzajemnie uwarunkowane, dokonano redukcji równań regresji przy wykorzystaniu metody regresji krokowej wstecznej. W jej efekcie otrzymano jedynie cztery statystycznie znaczące modele regresji (tab. 22).
Stwierdzono, że większość istotnych powiązań wystąpiła pomiędzy zadaniami wymagającymi maksymalnej aktywności oraz ich odpowiednikami, w których niezbędna była największa dokładność, czyli takimi, w których dążono do optymalizowania przejawów motoryczności. Konieczność optymalizacji była spowodowana wymogami przejawienia precyzyjnie określonej, wynoszącej połowę maksymalnych możliwości, wartości określonej zdolności. Za kondycyjne przejawy motoryczności uwarunkowane zdolnościami koordynacyjnymi można uznać (tab. 22):
maksymalną globalną siłę statyczną – z. nr 6;
częstotliwość ruchów lokalnych – z. nr 10;
czas reakcji prostej na bodziec optyczny – z. nr 4;
maksymalną lokalną siłę statyczną – z. nr 11.
Siła i częstotliwość ruchów były uzależnione od umiejętności precyzyjnego przejawiania połowy ich maksymalnej wartości (tab. 22). Natomiast czas reakcji prostej (z. nr 4) był związany z wielkością wykonania maksymalnego obrotu w wyskoku (z. nr 13). W każdym równaniu wartości BETA miały znak ujemny.
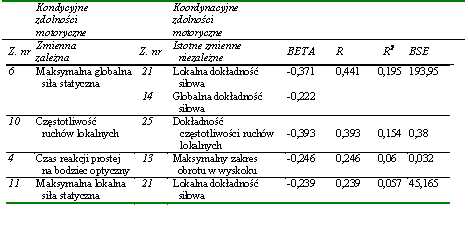
Przeprowadzono analizę skupień hierarchicznych, dokonując aglomeracji zmiennych, którymi były wszystkie kondycyjne i koordynacyjne zdolności motoryczne. Zmienne grupowano, stosując metodę Warda i posługując się odległościami euklidesowymi.
Na podstawie analizy skupień zdolności motorycznych można wnioskować o istnieniu bardzo dużych podobieństw pomiędzy niektórymi parami zadań motorycznych. Można do nich zaliczyć następujące, uszeregowane wg malejącego podobieństwa:
czas reakcji prostej (z. nr 4) i wysokość wyskoku pionowego (z. nr 5);
czasy uzyskania maksymalnej globalnej siły statycznej (z. nr 7) i maksymalnej lokalnej siły statycznej (z. nr 12);
częstotliwość ruchów globalnych (z. nr 3) i lokalnych (z. nr 10);
prędkość biegową (z. nr 1) i prędkość globalnego pojedynczego ruchu (prędkość wyprostu tułowia) (z. nr 2).
Kolejnymi parami, które cechowało znacznie mniejsze podobieństwo były:
dokładności wyskoku pionowego (z. nr 19) i obrotu w wyskoku (z. nr 20);
dokładność czasowa (z. nr 24) oraz dokładność częstotliwości ruchów lokalnych (z. nr 25) (ryc.7).
Analizując wyniki, stwierdzono, że zdecydowanie większym podobieństwem cechowały się zdolności kondycyjne niż koordynacyjne. Odległości pomiędzy kondycyjnymi zdolnościami motorycznymi zmieniały się w zakresie od 3,8 do 161,8. Jedyną zdolnością koordynacyjną, która wykazała bardzo duże podobieństwo do zdolności kondycyjnych, była lokalna dokładność siłowa (z. nr 21). Natomiast w bardzo interesujący sposób sklasyfikowano globalną (z. nr 6) i lokalną (z. nr 11) maksymalną siłę statyczną. Okazało się, że one zdecydowanie różniły się od większości analizowanych zdolności motorycznych (ryc. 7).
Wszystkie badane zdolności motoryczne zgrupowano w pięć zespołów cechujących się wewnętrznymi powiązaniami. Do pierwszego zaliczono zdolności wymagające umiejętności szybkiej orientacji w czasie i przestrzeni. Znalazły w niej miejsce takie parametry jak (ryc. 7):
czas reakcji prostej – z. nr 4;
wysokość wyskoku pionowego – z. nr 5;
czas uzyskania maksymalnej globalnej siły statycznej – z. nr 7;
czas uzyskania maksymalnej lokalnej siły statycznej – z. nr 12.
Odległości w obrębie tej grupy zdolności mieściły się w zakresie do 8,7 jednostki euklidesowej. Dowodzi to niespecyficznego, a zatem podobnego w zadaniach o globalnym i lokalnym oddziaływaniu, charakteru zdolności do przejawiania krótkiego czasu reakcji i szybkiego eksponowania siły oraz znacznej skoczności.
Do drugiej grupy zakwalifikowano szereg zadań motorycznych związanych z możliwościami szybkiego wykonywania pojedynczych ruchów o globalnym i lokalnym charakterze oraz realizacji ruchów z maksymalną częstotliwością (ryc. 7). Jednak podobieństwo pomiędzy tymi zdolnościami było mniejsze i wahało się od 15,4 do 80,0.
Obydwie grupy – pierwsza i druga – były podobne do siebie. Połączenie pomiędzy nimi nastąpiło w odległości równej 161,8 jednostek euklidesowych. Tę ostatnią wartość można uznać za miarę podobieństwa pomiędzy większością rozpatrywanych kondycyjnych zdolności motorycznych. W obydwu grupach znalazły się prawie wszystkie poza siłą statyczną. Natomiast jedyną koordynacyjną zdolnością motoryczną podobną do zdolności kondycyjnych była lokalna dokładność siłowa (z. nr 21).
W trzecim zespole zdolności motorycznych o stosunkowo dużym podobieństwie znalazły się zdolności koordynacyjne warunkujące precyzyjne wykonywanie ruchów o globalnym charakterze. Były to (ryc. 7):
dokładność wyskoku pionowego – z. nr 19;
dokładność obrotu w wyskoku – z. nr 20;
globalna dokładność siłowa – z. nr 14;
dokładność częstotliwości ruchów globalnych – z. nr 18.
Miary podobieństwa typowe dla tej grupy zdolności mieściły się w przedziale od 126,5 do 265,9 jednostek euklidesowych.
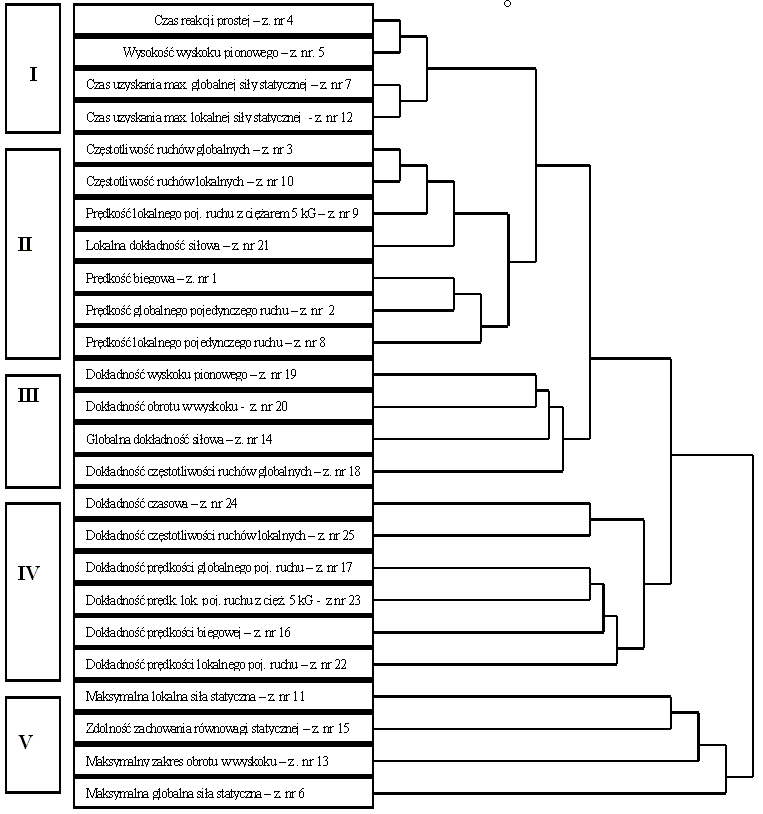
W kolejnej, czwartej, grupie znalazły się zadania motoryczne charakteryzujące zdolności precyzyjnego wykonywania ruchów, zarówno cyklicznych jak i acyklicznych, niezależnie od globalnego lub lokalnego przejawiania się aktywności mięśniowej (ryc. 7). Podobieństwo między nimi było zdecydowanie mniejsze, a jego miary, określane w jednostkach euklidesowych, mieściły się w przedziale od 335,5 do 753,8. W tej grupie znalazły się wyłącznie przejawy koordynacyjnych zdolności motorycznych.
Do piątej grupy zakwalifikowano zdolności o najmniejszym wzajemnym podobieństwie. Były nimi (ryc. 7):
maksymalna lokalna siła statyczna – z. nr 11;
zdolność zachowania równowagi statycznej – z. nr 15;
maksymalny zakres obrotu w wyskoku – z. nr 13;
maksymalna globalna siła statyczna – z. nr 6.
Odległości pomiędzy nimi oscylowały od 6606,3 do 44353,9 jednostek euklidesowych. W porównaniu z odległościami uzyskanymi w poprzednich grupach zdolności motorycznych, były to bardzo duże wartości. Można wnioskować, że te zdolności były niezależne. Interesujące, iż w składzie tej grupy były zarówno zdolności kondycyjne, jak i koordynacyjne. W przypadku zdolności koordynacyjnych były to wyłącznie zdolności globalne. Natomiast wśród kondycyjnych jedna miała charakter globalny, a druga lokalny. Wydaje się, że brak podobieństwa pomiędzy tymi zdolnościami sugeruje konieczność mierzenia każdej z nich podczas kompleksowej oceny zdolności motorycznych.
Podział zdolności motorycznych na kondycyjne i koordynacyjne jest w ujęciu niektórych autorów często kontrowersyjny. Ilustracją trudności w klasyfikacji może być umieszczenie zdolności szybkościowych jako hybrydowych, a zatem uwarunkowanych kondycyjnie i koordynacyjnie [Raczek 1993; Raczek i wsp. 2002].
Dążąc do uproszczenia tego zagadnienia, zaproponowano podział zadań ruchowych na maksymalne i optymalne. Podstawowym kryterium tego podziału stał się w tym przypadku zamierzony efekt zadania ruchowego. W maksymalnych przejawach motoryczności dąży się do uzyskania jak największego, maksymalnego efektu. Natomiast w optymalnych celem jest precyzyjne, dokładne, a zatem najbardziej zbliżone do zaplanowanego zrealizowanie zadania. Miarą możliwości jest odchylenie od wartości zadanej. W tym podziale nie ma żadnego znaczenia stopień złożoności ruchu, czyli podstawowe (często subiektywne) kryterium podziału na kondycyjne i koordynacyjne zadania motoryczne.
Na podstawie tak przyjętego podziału zgrupowano badane zadania motoryczne. Do maksymalnych zaliczono wszystkie zadania, w których celem było uzyskanie jak najlepszego (maksymalnego) wyniku, tj. największej siły, najszybszej reakcji, największej prędkości ruchu, największego obrotu w wyskoku czy największej wartości wyskoku pionowego oraz najmniejszej powierzchni pola rzutu pionowego ogólnego środka ciężkości ciała. Znalazły się tam zadania oznaczone numerami od 1 do 13 oraz zadanie nr 15 (tab. 10).
Natomiast do optymalnych przejawów zdolności motorycznych zakwalifikowano pozostałe zadania. Celem ich wykonania było osiągnięcie jak największej zgodności z wartością zadaną, mniejszą od maksymalnej. W realizowanych badaniach wynosiła ona połowę indywidualnej dla każdego badanego wartości maksymalnej. W tych zadaniach miarą była wartość popełnionego błędu, rozumiana jako odchylenie od wartości zadanej i podawana w %. Obliczano ją, stosując wzór nr 1. Możliwość wykonania zadania ruchowego z jak najmniejszym błędem uznano za zdolność optymalnego przejawienia zdolności motorycznych.
Porównując rezultaty typowe dla obydwu grup zdolności kondycyjnych stwierdzono jedynie nieliczne statystycznie znaczące współczynniki korelacji (tab. 23).
Taki charakter zależności oraz brak silnych związków wydaje się sugerować niewielkie uwarunkowania umiejętności precyzyjnego wykonania ruchu (zdolności do optymalnego przejawiania motoryczności) z możliwościami uzyskiwania w nim maksymalnej aktywności. Aby określić rzeczywiste powiązania pomiędzy obydwoma rodzajami aktywności ruchowej, ponownie posłużono się metodą analizy regresji wielokrotnej. W efekcie dokonanej redukcji stwierdzono, że niektóre zdolności o
nym charakterze aktywności, tj. wymagające dokładnego wykonywania ruchu, były uwarunkowane maksymalnymi możliwościami (tab. 24).
Na podstawie analizy rezultatów regresji wielokrotnej, w której zmiennymi zależnymi były zdolności motoryczne wymagające maksymalnej aktywności a zmiennymi niezależnymi ich przejawy, w których dążono do największej dokładności, można wnioskować o silnych wzajemnych uwarunkowaniach. Współczynniki korelacji wielokrotnej zmieniały się od 0,676 dla maksymalnego zakresu obrotu do 0,239 dla maksymalnej lokalnej siły statycznej (tab. 24).
Tabela 23 Zestawienie współczynników korelacji w parach zdolności motorycznych (maksymalne – optymalne) (n = 176)
 |
Nie było również wyraźnych zależności pomiędzy kondycyjnymi a koordynacyjnymi zdolnościami motorycznymi – określonymi w klasyczny sposób, tj. na podstawie oceny stopnia złożoności ruchu. Również zastosowanie podziału na wysiłki o maksymalnym i optymalnym charakterze aktywności mięśniowej doprowadziło do potwierdzenia tych zależności. Wszystkie miały ujemny charakter (tab. 24). Zatem większe możliwości w zakresie przejawiania maksymalnych wartości zdolności motorycznych warunkowały wyższy poziom umiejętności dokładnego, z mniejszym błędem, ich odtworzenia (tab. 25, 26, 27).
Nie stwierdzono natomiast żadnych znaczących uwarunkowań pomiędzy globalnymi a lokalnymi zdolnościami motorycznymi o optymalnym przejawianiu się.
Tabela 24 Charakterystyki statystycznie istotnych równań regresji wielokrotnej pomiędzy zdolnościami motorycznymi o maksymalnym i optymalnym charakterze aktywności (n = 176)
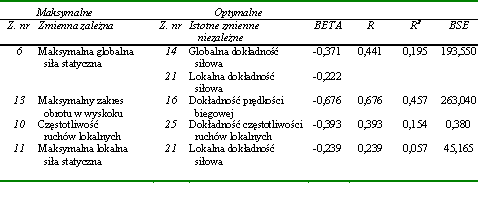 |
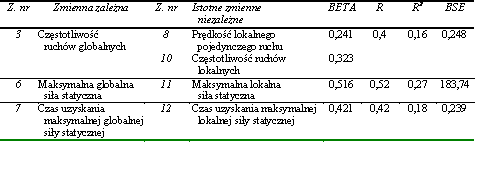


Statyczną siłę maksymalną zmierzono w zadaniach motorycznych wymagających globalnego oraz lokalnego jej przejawiania się. Określono też minimalny czas niezbędny do uzyskania największej wartości siły. Zastosowano także zadania motoryczne niezbędne do oceny dokładności siłowej poprzez pomiar umiejętności precyzyjnego osiągnięcia 50% maksymalnej globalnej i lokalnej siły statycznej. Również w tych zadaniach obliczano czasy konieczne do osiągnięcia zadanej wartości siły.
Stwierdzono, że czasy uzyskiwania maksymalnej siły globalnej i maksymalnej siły lokalnej były zbliżone i wynosiły odpowiednio 0,750 s i 0,794 s (ryc. 8, 9, 10). Wartości siły były oczywiście zdecydowanie różne. W przypadku siły globalnej jej średnia wartość wynosiła 1447,1 N (ryc. 8). Natomiast średnia wartość siły lokalnej ukształtowała się na poziomie 287,5 N (ryc. 9). Wystąpienie tego zjawiska dowodziło, że czas niezbędny do uzyskania maksymalnego napięcia mięśniowego i przejawienia największej siły był wartością niezależną od rodzaju ćwiczenia i ilości oraz rodzaju zaangażowanych grup mięśniowych. Przypuszczalnie może to być indywidualna charakterystyka osobnicza, w prezentowanych bowiem badaniach te wartości były znacznie zróżnicowane.
Wprowadzenie dodatkowego utrudnienia, jakim była konieczność uzyskania określonej (zadanej) siły – w pomiarze dokładności siłowej spowodowało bardzo wyraźne zmniejszenie dynamiki narastania siły w czasie. Wartości o połowę mniejsze osiągano w niewiele krótszym czasie.
Potwierdziło to wcześniej uzyskane rezultaty [Rynkiewicz, Starosta 2000]. Można zatem twierdzić, że w wyniku skomplikowania zadania ruchowego poprzez wprowadzenie kryterium dokładnego przejawienia siły, powstała konieczność stworzenia planu jego wykonania. Było to na tyle złożone, że znacząco wydłużało czas realizacji zadania. Interesujące, że zmniejszenie tempa przyrostu wielkości siły było szczególnie wyraźne w drugiej fazie jej zwiększania. W tej właśnie części zadania bardzo istotna stawała się precyzyjna kontrola przebiegu ruchu. Ważne wydaje się stwierdzenie, że spadek szybkości narastania siły był podobny w zadaniach o lokalnym i globalnym charakterze.
Rycina 8 Siła i czas jej narastania w pomiarach maksymalnej globalnej siły statycznej i globalnej dokładności siłowej (n = 176)


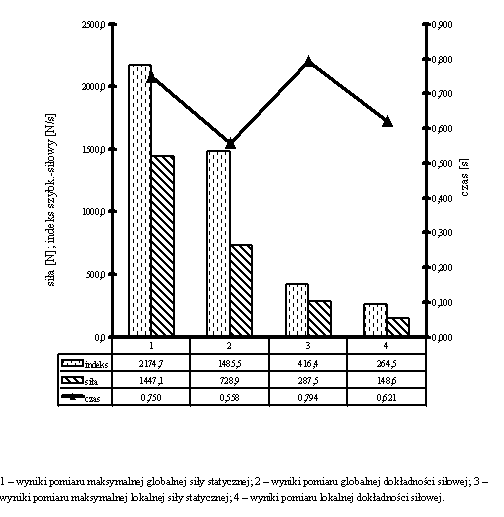
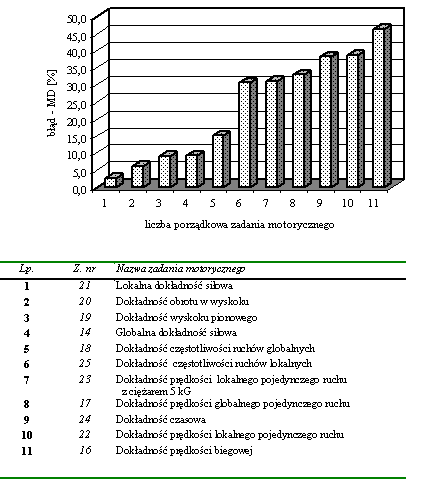
Istotnym problemem było określenie zależności pomiędzy rodzajem ruchu a możliwościami jego precyzyjnego dozowania. Dążąc do rozwiązania tego problemu, uszeregowano wyniki wszystkich zadań motorycznych wymagających dokładności. W trakcie analizy stwierdzono bardzo duże ich zróżnicowanie (ryc. 11).
Najmniejsze błędy były typowe dla wyników lokalnej dokładności siłowej (z. nr 21). Było to oczekiwanym rezultatem. Dokładność siłowa w zakresie ruchów lokalnych powinna być bardzo dobrze rozwinięta. Jej poziom warunkuje możliwości precyzyjnego wykonywania niezbyt obszernych ruchów, które są typowe dla życia codziennego. W dalszej kolejności znalazły się dokładność obrotu w wyskoku (z. nr 20) oraz dokładność wyskoku pionowego (z. nr 19) i globalna dokładność siłowa (z. nr 14).
Analiza istotności różnic pomiędzy średnimi arytmetycznymi wyników umożliwiła wyszukanie podobieństw jedynie pomiędzy dwoma ostatnimi zdolnościami (tab. 28). Pomimo to wydaje się, że najbardziej precyzyjnymi działaniami motorycznymi były wymagające przejawiania siły i mocy w pojedynczych aktach ruchowych oraz w zadaniach motorycznych wykonywanych w warunkach statyki.
W drugiej grupie zadań motorycznych, o mniejszych możliwościach dokładnego ich wykonywania, znalazły się wymagające osiągania znacznych prędkości pojedynczego ruchu bez obciążenia albo z niewielką jego wartością. Do niej zakwalifikowano też zadania, w których ważna była zdolność do przejawiania maksymalnej częstotliwości. Charakterystyczne, że podobnie jak w poprzedniej grupie, również ta umiejętność nie zależała od zakresu ruchu. Była podobna dla zadań o lokalnym i globalnym charakterze. Poziomem dokładności typowej dla tej grupy charakteryzowała się także zdolność dokładnego odtworzenia zadanego odcinka czasu, który dla badanych wynosił 1,000 s (ryc. 11). Większość zdolności z tej grupy miała zbliżone wartości. W zdecydowanej większości zadań nie znaleziono podstaw do wnioskowania o istnieniu statystycznie znaczących różnic pomiędzy średnimi arytmetycznymi zadań motorycznych zastosowanych do oceny poszczególnych zdolności.
Tabela 28 . Zestawienie istotności różnic pomiędzy średnimi wartościami błędów w zadaniach optymalizacyjnych (n = 176).
Tabela 28 Zestawienie istotności różnic pomiędzy średnimi wartościami błędów w zadaniach optymalizacyjnych (n = 176)
 |
Umiejętność uzyskania połowy prędkości maksymalnej na dystansie 5 m była u badanych mężczyzn najsłabsza. Wyniki cechowały się średnio błędem wynoszącym ponad 45%. Przypuszczalnie jego uwarunkowań należy poszukiwać w stopniu złożoności ruchu, którego wykonanie wymagało zaangażowania znacznej ilości dużych grup mięśniowych. Musiały one pracować z wysoką częstotliwością. Ponadto w zadaniu, w którym konieczne było przemieszczenie całego ciała w przestrzeni, podstawowym czynnikiem dostarczającym danych do regulacji intensywności był wzrok.
W rozwoju motoryczności zdolność do symetrycznego jej przejawiania ma duże znaczenie. Jego rola była wielokrotnie podkreślana. Ze względu na rangę problemu podjęto go też w niniejszych badaniach. Dlatego porównano rezultaty osiągnięte w takich samych ćwiczeniach wykonywanych prawą i lewą kończyną bądź w prawą lub lewą stronę. Uzyskane dane przedstawiono w tabeli 29. Na podstawie analizy współczynników korelacji liniowej można wnioskować o istnieniu bardzo silnych powiązań pomiędzy badanymi zdolnościami. Ich wartości zmieniały się w zakresie od 0,189 do aż 0,961. Najniższy znaczący współczynnik korelacji odnotowano w przypadku związku pomiędzy zdolnościami do precyzyjnego odtworzenia połowy wartości obrotu w prawo lub w lewo. Natomiast największą jego wartość odnotowano pomiędzy wynikami maksymalnego obrotu w prawo i lewo (tab. 29). Pozostałe współczynniki miały wartości pośrednie. Większość była znacząca statystycznie. Jedynie w dwóch przypadkach nie odnotowano istotnych związków pomiędzy symetrycznymi przejawami zdolności motorycznych. Nie było znaczącej zależności między wartościami lokalnej dokładności siłowej oraz w zadaniach motorycznych zastosowanych do oceny dokładności przejawiania częstotliwości ruchów lokalnych wykonywanych prawą i lewą kończyną górną (tab. 29).
Także analiza istotności różnic pomiędzy średnimi arytmetycznymi doprowadziła do potwierdzenia powyżej opisanych związków. Większość średnich arytmetycznych w parach symetrycznie wykonywanych zadań była podobna. Nie stwierdzono istotnych różnic pomiędzy typowymi dla nich wartościami średnimi (tab. 29). Wyjątkami były zadania wymagające lokalnej aktywności prawej lub lewej kończyny górnej, takie jak:
prędkość lokalnego pojedynczego ruchu (z. nr 8);
prędkość lokalnego pojedynczego ruchu z dodatkowym ciężarem 5 kG (z. nr 9);
częstotliwość ruchów lokalnych (z. nr 10);
maksymalna lokalna siła statyczna (z. nr 11);
dokładność częstotliwości ruchów lokalnych (z. nr 25).
Rezultaty typowe dla zadań motorycznych wykonywanych prawą i lewą kończyną przedstawiono w formie wartości względnych. Za 100% przyjęto wynik uzyskany w ćwiczeniach prawą kończyną bądź wykonywanych w prawą stronę. Do niego odnoszono rezultaty osiągane przy wykorzystaniu drugiej kończyny lub realizowanych w lewą stronę. Na tej podstawie można ponownie stwierdzić, że niewielką przewagę miały zadania motoryczne wykonywane w prawo (ryc. 12).
Porównanie średnich arytmetycznych i analiza różnic pomiędzy nimi wydaje się niewystarczająco skutecznym narzędziem do badania stopnia symetryzacji. Nie uwzględnia się bowiem rozproszenia danych, którego miarą jest średnie odchylenie standardowe. Dlatego zaproponowano zastosowanie wskaźnika symetryzacji, który umożliwia określenie różnic pomiędzy dwoma czynnościami ruchowymi, z uwzględnieniem wartości średnich arytmetycznych i średnich odchyleń standardowych [Szopa i wsp. 1985]. Symetrycznie przejawiane zdolności motoryczne uszeregowano zgodnie z narastaniem wskaźnika symetryzacji, czyli odwrotnie do stopnia symetrii (ryc. 12)
Stwierdzono, że największe możliwości symetrycznego wykonania były typowe dla zadań motorycznych związanych z dokładnością prędkości pojedynczego ruchu wykonywanego bez dodatkowego obciążenia (z. nr 22) i z koniecznością pokonania ciężaru 5 kG (z. nr 23). Na podobnym poziomie ułożyły się wartości wskaźnika symetryzacji typowe dla czasu uzyskania zadanej siły w pomiarze lokalnej dokładności siłowej (z. nr 27) oraz maksymalny zakres obrotu w wyskoku (z. nr 13) i lokalna dokładność siłowa (z. nr 21) a także czas uzyskania maksymalnej lokalnej siły statycznej (z. nr 12) (ryc. 12). Pozostałe zdolności motoryczne charakteryzowały się większymi wartościami wskaźnika symetryzacji. Natomiast zdecydowanie najsłabsze wyniki typowe były dla częstotliwości ruchów lokalnych (z. nr 10) (ryc. 12). Porównanie wskaźników symetryzacji sugerowało istnienie wyższego poziomu zdolności motorycznych prawej strony lub prawej kończyny.
Konieczność precyzyjnego wykonania ruchów kończyną górną była połączona z mniejszą asymetrią niż dążenie do uzyskania w nich maksymalnych wartości. Przypuszczalnie było to związane z lepszym wyćwiczeniem kończyny dominującej. Wprowadzenie elementu nowego, w przypadku wymagań dotyczących dokładności, wyeliminowało wpływ wytrenowania nabytego poprzez częstsze jej używanie, które nałożyło się na uwarunkowaną genetycznie stronność.
To ostatnie zjawisko nie dotyczyło zadań ruchowych o globalnym charakterze. W tym przypadku maksymalny obrót w wyskoku cechowała jedynie nieco większa symetryczność niż jego optymalne (dokładne) przejawienie (ryc. 12). Można przypuszczać, że globalny lub lokalny charakter aktywności mięśniowej nie różnicował pod względem asymetrii czynnościowej w zakresie badanych zdolności motorycznych. Obydwa zadania motoryczne o globalnym charakterze mieściły się w połowie zestawienia.
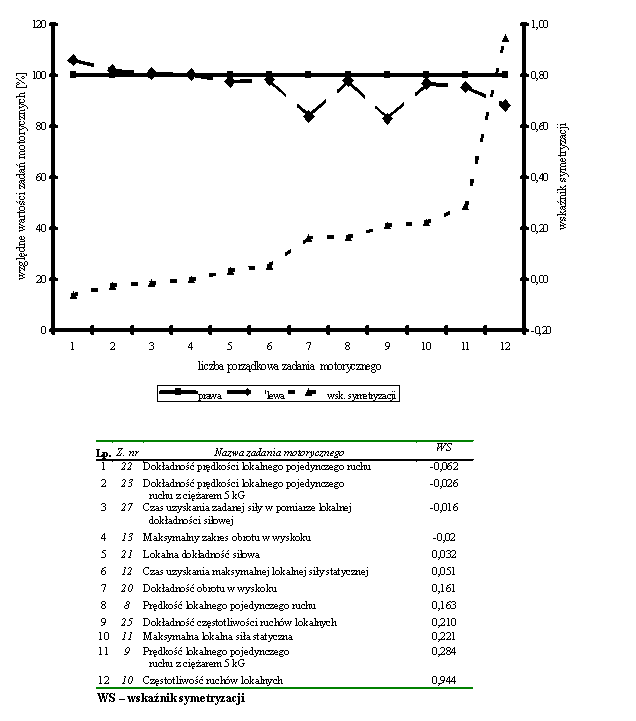
Porównanie rezultatów osiągniętych przez kobiety i mężczyzn w zadaniach motorycznych zastosowanych do analizy kondycyjnych i koordynacyjnych zdolności motorycznych upoważniło do wnioskowania o istnieniu szeregu znaczących różnic (tab. 30). Większość wystąpiła w obrębie zdolności kondycyjnych.
Szczególnie silne rozbieżności pomiędzy wynikami typowymi dla przedstawicieli obydwu płci odnotowano w zakresie globalnych kondycyjnych zdolności motorycznych. Znaczących różnic nie było jedynie w przypadku dwóch zadań, którymi były czas reakcji prostej (z. nr 4) oraz czas uzyskania maksymalnej globalnej siły statycznej (z. nr 7), co ukazano w tabeli 30.
Rozpatrując wyniki typowe dla lokalnych zdolności kondycyjnych, stwierdzono, że istotne różnice wystąpiły w prędkości pojedynczego ruchu kończyną górną (z. nr 8) oraz przejawianej maksymalnej lokalnej siły statycznej (z. nr 11). W każdej z tych zdolności mężczyźni przejawiali zdecydowanie wyższe wartości średnie (tab. 30).
Kobiety cechowała znacznie mniejsza siła maksymalna i zdolność do jej przejawiania w dynamicznych wysiłkach. Dlatego miały one mniejsze wartości prędkości biegowej (z. nr 1), prędkości globalnego pojedynczego ruchu (z. nr 2), mniejszą częstotliwość ruchów globalnych (z. nr 3) oraz niższe wartości wyskoku pionowego (z. nr 5) (tab. 30).
Średnie wartości uzyskane w zadaniach motorycznych związanych z koniecznością przejawienia reaktywności i ruchliwości układu nerwowego były zbliżone w obydwu grupach. Dlatego nie stwierdzono różnic w czasie reakcji prostej na bodźce optyczne (z. nr 4). Podobne były też wartości czasu osiągania maksymalnej globalnej i lokalnej siły statycznej (z. nr 7 i z. nr 12) oraz częstotliwość ruchów lokalnych (z. nr 10). Interesujące, że kobiety miały większe wartości w zadaniu polegającym na jak najszybszym wykonaniu lokalnego pojedynczego ruchu z obciążeniem ciężarem 5 kG (z. nr 9). Przyczyną była prawdopodobnie zbyt mała dla mężczyzn wartość ciężaru. Jego zastosowanie powodowało nadmiernie silne pobudzenie, które ograniczało zdolność do szybkiego wykonania ruchu. Prędkość pojedynczego ruchu z ciężarem wymaga rozwinięcia mocy. Uważa się, że optymalne warunki do jej przejawienia występują przy 1/3 maksymalnej prędkości oraz ½ maksymalnej siły [Zaciorski 1970; Trzaskoma, Trzaskoma 2001]. Dla kobiet odznaczających się mniejszymi możliwościami w zakresie maksymalnej siły statycznej zastosowane warunki były bliższe najlepszym do przejawiania się maksymalnej mocy (tab. 30).
W zakresie koordynacyjnych zdolności motorycznych stwierdzono niewiele różnic w wynikach typowych dla kobiet i mężczyzn. Zróżnicowanie statystycznie istotne odnotowano jedynie w trzech zadaniach motorycznych. I tak, kobiety miały lepsze wyniki w zakresie maksymalnego obrotu (z. nr 13). Jest to interesujące, ponieważ stwierdzono, że wartość obrotu w wyskoku zależała od zdolności do szybkiej reakcji oraz od umiejętności kontrolowania położenia części ciała w przestrzeni. Okazuje się, że przy podobnych umiejętnościach w tym zakresie kobiety potrafiły wykonać większy obrót w wyskoku. Przypuszczalnie za jedną z przyczyn można uznać mniejszy moment obrotowy, typowy dla kobiet, spowodowany mniejszą masą ciała u badanych kobiet niż u mężczyzn. Badane kobiety uzyskiwały niższe wyniki od mężczyzn w umiejętności dokładnego odtworzenia prędkości globalnego pojedynczego ruchu (wyprostu tułowia) i lokalnej dokładności siłowej.
Tabela 30 Istotności różnic w kondycyjnych i koordynacyjnych zdolnościach motorycznych u mężczyzn (n = 176) i kobiet (n = 84)
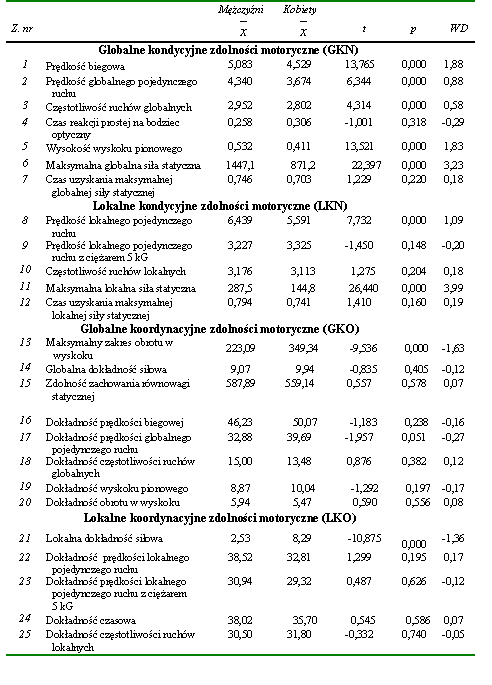 |
Ponieważ typowe dla nich były mniejsze wartości prędkości globalnego pojedynczego ruchu i maksymalnej siły statycznej – globalnej i lokalnej, można sądzić, że o dokładności w przejawianiu tych zdolności decydowała składowa siłowa, a nie koordynacyjna. Potwierdzeniem tego może być uprzednio stwierdzona ujemna zależność pomiędzy maksymalnymi a optymalnymi wartościami zdolności motorycznych. Natomiast pozostałe zdolności koordynacyjne określone przy wykorzystaniu zastosowanych zadań motorycznych były podobne u kobiet i mężczyzn (tab. 30).
Wydaje się, że zróżnicowanie płciowe było typowe głównie dla zdolności kondycyjnych. Największe różnice pomiędzy przedstawicielami obydwu płci odnotowano w zakresie globalnych zdolności kondycyjnych, wymagających przejawiania siły i mocy (ryc.13, ryc. 14).
Rycina 13 Względne wartości zdolności motorycznych w zespołach kobiet (n = 84) i mężczyzn (n = 176) – kondycyjne zdolności motoryczne

Rycina 14 Względne wartości zdolności motorycznych w zespołach kobiet (n = 84) i mężczyzn (n = 176) - koordynacyjne zdolności motoryczne

Natomiast możliwości precyzyjnego dozowania przejawów motoryczności, oceniane na podstawie wartości błędu od wartości zadanej, na ogół były podobne w przypadku zarówno kobiet jak i mężczyzn (tab. 31). Można wnioskować, że kobiety przy mniejszych możliwościach w zakresie przejawiania maksymalnych wartości cechowały podobne do mężczyzn zdolności optymalnego, a zatem dokładnego osiągania określonych wartości (tab. 31).
Nie stwierdzono także wyraźnych różnic pomiędzy zdolnością do przejawiania największej dokładności wykonania zadania ruchowego u kobiet i mężczyzn. Większość współczynników dymorfizmu w tych zadaniach motorycznych mieściła się w przedziale od -0,27 do 0,12 (tab. 31). Jedynym wyjątkiem były możliwości osiągania lokalnej dokładności siłowej (z. nr 21). W tym zadaniu badane kobiety przejawiły znacznie mniejsze możliwości. Wskaźnik dymorfizmu osiągnął wartość równą -1,36 (tab. 31). Wydaje się, że przyczyn należy upatrywać w zakresie umiejętności ćwiczonych w rozmaitych czynnościach życiowych i sportowych, które są różne u kobiet i mężczyzn.

Spis treści
- 7.1. Związki pomiędzy zdolnościami motorycznymi
- 7.2. Globalne a lokalne zdolności motoryczne
- 7.3. Kondycyjne a koordynacyjne zdolności motoryczne z uwzględnieniem ich globalnych i lokalnych przejawów
- 7.4. Maksymalne i optymalne przejawy globalnych i lokalnych zdolności motorycznych
- 7.5. Symetria w zakresie przejawiania globalnych i lokalnych zdolności motorycznych
- 7.6. Zróżnicowanie płciowe w zakresie globalnych i lokalnych przejawów zdolności motorycznych
Przed przystąpieniem do weryfikacji pierwszych dwóch hipotez, dotyczących relacji pomiędzy globalnymi i lokalnymi przejawami kondycyjnych i koordynacyjnych zdolności motorycznych, konieczne było określenie powiązań występujących w obrębie każdej grupy zdolności motorycznych. W badaniach własnych zdolności motoryczne podzielono w następujący sposób:
globalne zdolności kondycyjne – GKN;
lokalne zdolności kondycyjne – LKN;
globalne zdolności koordynacyjne – GKO;
lokalne zdolności koordynacyjne – LKO.
Ustalenie związków pomiędzy zdolnościami motorycznymi jest istotnym zagadnieniem ze względu na konieczność dokonania wyboru najbardziej znaczących z nich dla kontroli poziomu wytrenowania lub oceny możliwości wykonywania pracy [Szopa i wsp. 1999; Wachowski, Strzelczyk 1999]. Ponadto w wychowaniu fizycznym i sporcie niezbędne jest posiadanie informacji o wzajemnych powiązaniach i oddziaływaniu kształtowanych zdolności. Wyniki w zadaniach motorycznych wykorzystanych do analizy zdolności motorycznych w badaniach własnych były względnie niezależne, szczególnie w zakresie ich lokalnych przejawów. Jednak jednorodny materiał badawczy oraz niskie wartości współczynników korelacji sugerują konieczność zachowania ostrożności w analizie uzyskanych wyników.
Na podstawie uzyskanych rezultatów badań własnych nie znaleziono podstaw do wnioskowania o związkach globalnej i lokalnej maksymalnej siły statycznej z prędkością biegową. Siła jest zdolnością bardzo zróżnicowaną. Może być rozwijana w zadaniach ruchowych statycznych i dynamicznych. Natomiast ze względu na wielkość oporu koniecznego do pokonania może przejawiać się w różnych odmianach: pasywnej, aktywnej, wolnej, szybkiej i eksplozywnej [Zaciorski 1970; Ważny 1977; Naglak 1979; Trzaskoma, Trzaskoma 2001]. W badaniach własnych stosowano tylko pomiary w statyce. Zdolności siłowe są uwarunkowane przekrojem fizjologicznym mięśni, liczbą i stopniem unerwienia jednostek motorycznych, proporcjami pomiędzy włóknami mięśniowymi szybko- i wolnokurczliwymi oraz wskaźnikami budowy ciała związanymi głównie z proporcjami dźwigni kostnych [Szopa 1993; Trzaskoma, Trzaskoma 2001]. Bardzo ważna jest też możliwość uwalniania energii z rozpadu fosfokreatyny [Popinigis 1997].
Szybkość jest zdolnością zdeterminowaną głównie przez siłę i ruchliwość układu nerwowego, czyli indywidualnie względnie stałe wartości [Zaciorski 1970; Sozański, Witczak 1981; Szopa 1993]. Natomiast pozostałe uwarunkowania są podobne do typowych dla siły [Zaciorski 1970; Szopa 1993; Popinigis 1997]. Przejawienie szybkości wymaga wytworzenia znacznej energii w krótkim czasie, ale stawia też bardzo wysokie wymagania wobec sprawności układu nerwowego, szczególnie w zakresie koordynacji nerwowo-mięśniowej oraz koordynacji wewnątrzmięśniowej. Ponieważ są to bardzo zróżnicowane mechanizmy, dlatego przejawianie się szybkości może zależeć od rodzaju ruchu, szczególnie od jego cykliczności. Ponieważ przejawienie szybkości wymaga zarówno znacznego potencjału energetycznego, jak i sprawnego układu nerwowego, dlatego w efekcie wzrostu siły, nie popartego odpowiednim działaniem na poprawę koordynacji nerwowo-mięśniowej, może pogorszyć się płynność ruchów i ich rytm. Zaburzeniom może ulec koordynacja ruchowa [Zaciorski 1970; Dumitrescu 1975; Kruczalak 1979; Osiński 1977]. Wydaje się nawet, że zależność pomiędzy siłą statyczną a prędkością nie jest liniowa. Nadmierne zwiększenie siły prowadzi do pogorszenia możliwości szybkościowych [Wachowski i wsp. 1976; Osiński 1977; Sozański, Witczak 1981].
Stwierdzono istotną statystycznie zależność pomiędzy prędkością wyprostu (prostowania tułowia), częstotliwością przeskoków i pośrednio prędkością biegową oraz czasem narastania globalnej maksymalnej siły statycznej. Natomiast wyniki pomiaru wysokości pionowego wyskoku, czyli sprawdzianu często wykorzystywanego do pośredniej oceny mocy, nie korelowały z tymi zdolnościami oraz z globalną siłą statyczną. Było to nieoczekiwane zjawisko [Osiński 1988a]. Istnieją uzasadnienia zależności wyskoku i prędkości biegowej od dyspozycji siłowo-szybkościowych oraz prędkości prostowania tułowia [Ślężyński, Mynarski 1973; Osiński 1977; Popow, Stepanenko 1978]. Prawdopodobnie prędkość wyprostu w ruchu globalnym jest zdolnością związaną z szybkością narastania siły. Dopiero jej znaczny poziom w połączeniu z umiejętnością wykonywania ruchów o wysokiej częstotliwości warunkuje prędkość lokomocyjną oraz wysokość wyskoku. Nie znaleziono podstaw do wnioskowania o wpływie zdolności do szybkiej reakcji na poziom badanych przejawów kondycyjnych zdolności motorycznych, tj. szybkości, siły i mocy. W każdym ze stosowanych w badaniach własnych zadaniu motorycznym ćwiczący dowolnie wybierali moment startu. Mieli wystarczająco dużo czasu na koncentrację i opracowanie planu realizacji ruchu. Było to zgodne z wynikami innych badań [Henry 1960; Henry, Whitley 1960; Geblewiczowa 1965, 1968; Zaciorski 1970; Osiński i wsp. 1978; Sozański, Witczak 1981]. Przedstawione relacje wydają się dotyczyć głównie zadań o globalnym charakterze aktywności mięśniowej. Lokalne kondycyjne zdolności motoryczne prawdopodobnie są bardziej specyficzne, a zatem mniej wzajemnie uwarunkowane. Natomiast pozostałe zdolności motoryczne można uznać za względnie niezależne.
Podobne rezultaty uzyskała Wyżnikiewicz-Kopp [1992]. Badając zależności w obrębie zdolności kondycyjnych (bateria testów EUROFIT) oraz koordynacyjnych zdolności motorycznych, stwierdzono więcej powiązań wśród tych pierwszych. Uzasadniono to wspólnym podłożem biologicznym kondycyjnych zdolności motorycznych i wysoką specyficznością koordynacyjnych zdolności motorycznych [Wyżnikiewicz-Kopp 1992].
Zgodnie z poglądami innych autorów, u podstaw wszystkich działań ruchowych leżą procesy informacyjne i energetyczne. Natomiast w konkretnych zadaniach motorycznych rozmaity jest ich udział [Raczek i wsp. 2002]. Wydaje się, że siłowe i szybkościowo-siłowe zdolności motoryczne były uwarunkowane głównie możliwościami energetycznymi, z natury mało specyficznymi. Natomiast do uzyskania wysokich wartości w bardziej skomplikowanych zadaniach ruchowych, które charakteryzowały wyższe wymagania w stosunku do zdolności koordynacyjnych, niezbędne było posiadanie silnego i ruchliwego układu nerwowego, którego możliwości były specyficzne, a zatem rozmaite dla poszczególnych zadań motorycznych. Można przyjąć, że koordynacyjne zdolności motoryczne mają bardziej złożoną strukturę, a jej poszczególne składowe są względnie niezależne. O zakresie rozbieżności w ocenie struktury koordynacyjnych zdolności motorycznych świadczy wydzielanie przez różnych autorów od 2–3 ogólnych do nawet 20 specjalnych i specyficznych koordynacyjnych zdolności motorycznych [Ljach 1987, 1989, 1990, 2001; Mynarski 1986, 2000; Combee 1970; Filipowicz, Turewski 1977; Fleischman 1972; Guilford 1958; Hirtz 1977, 1985; Ilin 1976; Mĕkota 1982; Wiszniakow 1993].
Istniejące zróżnicowanie poglądów sugeruje konieczność zachowania ostrożności we wnioskowaniu. W badaniach własnych nie zbadano trafności zadań motorycznych zastosowanych do pomiaru koordynacyjnych zdolności motorycznych. Ponadto charakter niektórych zadań testowych spowodował stosunkowo niskie wartości miar rzetelności. Z tych powodów interpretacja wyników badań własnych nie może być uznawana za całkowicie pewną.
Określenie powiązań pomiędzy globalnymi a lokalnymi przejawami motoryki człowieka stanowi bardzo istotne zagadnienie dla rozszerzenia teoretycznych podstaw nauki o ruchu. Jest to dotychczas stosunkowo mało zbadany aspekt nauki o ruchu [Hirtz 1985; Mynarski 2000; Starosta 2001a]. Równocześnie w praktyce sportowej często wyznaczano cele i sposoby treningu siły z podziałem na jej globalne i lokalne przejawy [Zaciorski 1970; Trzaskoma, Trzaskoma 2001; Waśkiewicz 2002]. Większość danych dotyczyła głównie lokalnych i globalnych przejawów koordynacji ruchowej [Rutkowska-Kucharska 1982; Rutkowska-Kucharska i wsp. 1990; Ljach 1989, 1990, 2001]. Bober, Golema, Rutkowska-Kucharska [1990] zdefiniowali ruchy lokalne jako te, w których pracują mięśnie działające na jeden staw, z jednym stopniem swobody. Natomiast ruchy wymagające zaangażowania większej liczby mięśni oraz odbywające się przy większej liczbie stopni swobody klasyfikowano jako globalne. Identyczny podział ruchów przyjęto w badaniach własnych.
Analizując powiązania pomiędzy globalnymi a lokalnymi przejawami zdolności motorycznych, stwierdzono zależności zróżnicowane pod względem siły i charakteru. Zdolności kondycyjne były często skorelowane, zwłaszcza ich globalne przejawy. Natomiast koordynacyjne były prawie całkowicie niezależne. Jednak interpretacja tych związków wymaga zachowania pewnej dozy ostrożności ze względu na niezbyt wysokie wartości współczynników korelacji.
Wykorzystanie do analizy wyników metody redukcji regresji wielokrotnej umożliwiło bardziej dokładne określenie powiązań występujących pomiędzy globalnymi a lokalnymi zdolnościami motorycznymi. W obrębie zadań motorycznych wykorzystanych do analizy zdolności kondycyjnych, po pełnej redukcji równania regresji wielokrotnej, pozostawały zależności pomiędzy maksymalną globalną i lokalną siłą statyczną, minimalnymi czasami koniecznymi do osiągania ich największych wartości oraz korelacja pomiędzy częstotliwością ruchów globalnych i lokalnych. Istotne znaczenie miała też prędkość pojedynczego ruchu o lokalnym charakterze – warunkowała ona częstotliwość ruchów globalnych.
Rozpatrując współczynniki korelacji zaobserwowano jedynie niewielką liczbę zależności między globalnymi a lokalnymi zdolnościami koordynacyjnymi. Natomiast zastosowanie analizy regresji upoważniło do wnioskowania o całkowitym braku statystycznie istotnych powiązań pomiędzy globalnymi a lokalnymi przejawami tych zdolności. Nie stwierdzono żadnych znaczących zmiennych niezależnych, tj. takich, które pozostawały po zakończeniu redukcji składowych równań. Można twierdzić, że w obrębie koordynacyjnych zdolności motorycznych nie było wyraźnych związków pomiędzy ich globalnymi i lokalnymi przejawami (ryc. 15). Wydaje się, że brak związków jest uzależniony od typowego dla człowieka zróżnicowania mechanizmów sterowania ruchami.
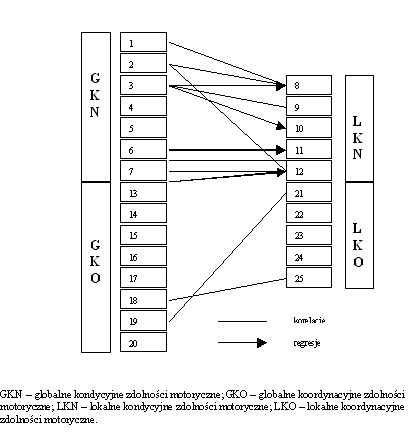
Także analiza skupień upoważniła do stwierdzenia, że większym podobieństwem charakteryzowały się zdolności globalne. Równocześnie zaobserwowano zdolności zupełnie różne od pozostałych. Do nich należały: siła, maksymalny obrót w wyskoku i zdolność zachowania równowagi. Wydaje się, że przy kompleksowym pomiarze zdolności motorycznych należy stosować je wszystkie.
Pierwsza hipoteza znalazła potwierdzenie w analizie wyników przeprowadzonych badań. Stwierdzono, że lokalne i globalne koordynacyjne zdolności motoryczne były niezależne. Do podobnych wniosków doszedł Ljach [1990], który u dzieci (chłopców i dziewcząt) w wieku od 7 do 17 lat analizował około 5000 korelacji pomiędzy heterogennymi, elementarnymi (lokalnymi) i globalnymi wskaźnikami koordynacyjnych zdolności motorycznych. W tych badaniach aż w 82–97% przypadków nie stwierdził statystycznie znaczących współczynników korelacji [Ljach 1990].
Symptomatyczne, że większość autorów zajmujących się powiązaniami pomiędzy globalnymi i lokalnymi zdolnościami motorycznymi ograniczyła się jedynie do ich określenia, natomiast nie analizowała przyczyn takiego stanu. Nie podjęto też próby teoretycznego uzasadnienia tego zjawiska [Ljach 1990].
Dążąc do realizacji pierwszego celu pracy, którym było określenie zależności między lokalnymi a globalnymi przejawami kondycyjnych i koordynacyjnych zdolności motorycznych, stwierdzono w badaniach własnych jedynie częściowe powiązania w obrębie lokalnych i globalnych przejawów kondycyjnych zdolności motorycznych. Natomiast lokalne i globalne przejawianie się koordynacyjnych zdolności motorycznych uznano za niezależne. Wydaje się, że przyczyn należy upatrywać w różnych mechanizmach warunkujących globalne oraz lokalne kondycyjne i koordynacyjne zdolności motoryczne. W zadaniach o globalnym i lokalnym charakterze aktywności podobny jest potencjał energetyczny, szybkość pobudzania mięśni oraz ruchliwość układu nerwowego. Zatem takie podstawy zdolności motorycznych są niespecyficzne. Za podstawowe różnice w zakresie lokalnych i globalnych przejawów zdolności motorycznych można uznać uwarunkowania informacyjne związane z posiadaniem określonych programów (wzorców) wykonywania ruchów lub tworzeniem nowych na bazie istniejącego potencjału. Te uwarunkowania zdolności motorycznych można uznać za specyficzne. Mogą one być odmienne dla globalnych i lokalnych przejawów zdolności motorycznych.
Podstawy energetyczne przejawiania zdolności do wykonywania pracy rozpatrywano na poziomie globalnym lub lokalnym [Zaciorski 1970; Naglak 1979]. Obecnie dominuje pogląd o ich niespecyficznym charakterze, szczególnie w przypadku globalnych uwarunkowań sprawności mechanizmów energetycznych glikolizy tlenowej i beztlenowej. Uważa się też, że możliwości przejawiania znacznej koordynacji nerwowo-mięśniowej zależą głównie od indywidualnych właściwości osobniczych, uwarunkowanych określonym poziomem sprawności układu nerwowego oraz od posiadanego zasobu nawyków ruchowych. Jest to szeroko wykorzystywane w praktyce wychowania fizycznego i sportu przy doskonaleniu kondycyjnych zdolności motorycznych oraz różnych przejawów koordynacji ruchowej [Zaciorski 1970; Szopa i wsp. 1996; Prus 2000; Raczek i wsp. 2002]. Możliwości wytrenowania kondycyjnych zdolności motorycznych są niekwestionowane. Także w zakresie koordynacyjnych zdolności motorycznych istnieją znaczne możliwości poprawy w efekcie oddziaływania treningowego [Zimmermann 1982, 1983, 1986; Szczepanik 1993; Szopa 1993; Szopa i wsp. 1996; Prus, Szopa 1997; Prus 2000].
Istotnym problemem postawionym w drugiej hipotezie było określenie relacji występującej pomiędzy energetycznymi (kondycyjnymi) a informacyjnymi (koordynacyjnymi) zdolnościami motorycznymi. Niektórzy autorzy wskazywali na wagę tego zagadnienia [Hirtz, Starosta 1990; Ljach 1990; Wyżnikiewicz-Kopp 1992; Starosta 1995a]. Na podstawie znanych z dostępnego piśmiennictwa rezultatów badań można wnioskować, że były one zmienne i niedostatecznie rozpoznane [Blume 1981; Hirtz, Starosta 1990; 1995a]. Dlatego analizując wyniki badań własnych postanowiono uwzględnić to zagadnienie w aspekcie globalnego i lokalnego charakteru przejawiania się zdolności motorycznych. Na podstawie analizy współczynników korelacji uzyskanych w badaniach własnych nie można jednoznacznie wnioskować o silnych koordynacyjnych uwarunkowaniach zdolności kondycyjnych. Niemniej jednak takie zależności zaistniały. Były one silniejsze w przypadku zdolności motorycznych o globalnym charakterze. Chociaż także w tym przypadku miary zależności nie były zbyt wysokie (ryc. 16). Stanowiło to jedynie częściowe potwierdzenie drugiej hipotezy.
Rycina 16 Zależności pomiędzy globalnymi i lokalnymi przejawami kondycyjnych i koordynacyjnych zdolności motorycznych (n = 176)
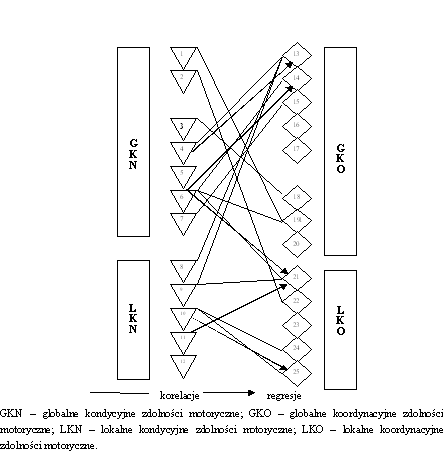
W badaniach własnych analizowano globalne i lokalne przejawy kondycyjnych zdolności motorycznych o podłożu szybkościowym. Z rezultatów dotychczasowych badań wynikał brak uwarunkowań koordynacyjnych zdolności motorycznych różnymi przejawami wytrzymałości [Havli ek 1972; Mĕkota 1982; Raczek 1987; Ljach 1990; Raczek i wsp. 1998, 2002]. Natomiast opierając się na wynikach uzyskanych przez Ljacha [1990], można było wnioskować o związkach pomiędzy koordynacyjnymi zdolnościami motorycznymi a zdolnościami szybkościowymi i szybkościowo-siłowymi. Istotne korelacje stwierdzono u 30 % badanych [Ljach 1990]. Nie doszukano się też istotnych współzależności koordynacyjnych zdolności motorycznych ze wskaźnikami siły maksymalnej. Większość współczynników korelacji nie przekraczała 0,30 [Turewski 1993; Fleischman 1964; Havli ek 1972; Vilkner 1986; Ljach 1991b]. Ustalono też, że w większości przypadków koordynacyjne zdolności motoryczne nie były związane z gibkością [Mĕkota 1982; Filipowicz, Turewski 1977]. Stwierdzono natomiast, że koordynacyjne zdolności motoryczne były bardziej skorelowane z kondycyjnymi zdolnościami motorycznymi o charakterze szybkościowym i siłowo-szybkościowym [Ljach 1976, 1990; Turewski 1993; Mĕkota 1982; Vilkner 1989; Raczek i wsp. 1998, 2002].
Zbliżone rezultaty uzyskała w swoich badaniach Wyżnikiewicz-Kopp [1992], która również nie znalazła zbyt wielu powiązań pomiędzy kondycyjnymi (badanymi zestawem testów EUROFIT) a koordynacyjnymi zdolnościami motorycznymi.
Należy podkreślić, że zdolności koordynacyjne są uwarunkowane głównie możliwościami centralnego i obwodowego układu nerwowego [Bernstein 1966, 1991; Starosta 1990; Raczek i wsp. 2002]. Zależą od mechanizmów wyznaczających możliwości sterowania ruchem i jego regulacją. Natomiast zdolności kondycyjne determinuje przede wszystkim potencjał energetyczny. Niezależnie od tego czy przyjmiemy ten pogląd, czy też teorię o jedności procesów informacyjnych i energetycznych [Krüger 1989; Vilkner 1989], zawsze przejawianie się zdolności motorycznych będzie wymagało równoczesnego zaangażowania jednych i drugich. Proporcje pomiędzy nimi będą zależały jedynie od celu działania [Raczek i wsp. 2002]. Zdolności motoryczne przejawiają się w konkretnych umiejętnościach ruchowych. Przy czym zdolności motoryczne stanowią ich ogólne uwarunkowanie, a umiejętności ruchowe powinny być traktowane jako specjalne podłoże dla konkretnych działań ruchowych [Raczek i wsp. 2002].
Wydaje się, że pogląd o jedności procesów energetycznych i informacyjnych jest bardziej racjonalny. Wykonanie jakiegokolwiek zadania ruchowego wymaga przejawienia energii. Natomiast sposób jej użycia jest zdeterminowany działaniem układu nerwowego. Jego możliwości zależą od współdziałania receptorów, dróg przewodzących bodźce oraz od sprawności ośrodków kojarzeniowych, w których tworzone są programy działania. Niebagatelne znaczenie ma też zasób posiadanych nawyków ruchowych.
W analizie powiązań pomiędzy kondycyjnymi a koordynacyjnymi zdolnościami motorycznymi wyodrębniono maksymalny obrót wokół podłużnej osi ciała, wykonywany w wyskoku, jako globalny sposób oceny zdolności koordynacyjnych. Ten zaproponowany przez Starostę test jest prosty w realizacji, a równocześnie umożliwia zebranie dokładnych wyników [Starosta 1978a]. Na podstawie analizy współczynników korelacji stwierdzono, że rezultaty tego testu zależały jedynie od zdolności przejawiania szybkiej reakcji oraz umiejętności wykonania szybkich ruchów. Były to jego podstawowe uwarunkowania kondycyjne. Należy podkreślić, że uzyskane współczynniki korelacji były niezbyt wysokie, chociaż znaczące statystycznie. Do zasadniczych determinant koordynacyjnych zaliczono natomiast dokładność w przemieszczaniu swojego ciała i zdolność zachowania równowagi. Kierując się wynikami analizy regresji wielokrotnej, można stwierdzić, że wartość maksymalnego obrotu w wyskoku zależała głównie od globalnych zdolności koordynacyjnych, szczególnie od dokładności odtworzenia połowy prędkości lokomocyjnej – czyli umiejętności dokładnego przemieszczania całego ciała w przestrzeni. Zdolności kondycyjne miały mniejsze znaczenie (niezależnie do globalnego lub lokalnego charakteru ich przejawiania się), a podstawową z nich była szybkość reakcji. Ujemne wartości współczynnika świadczyły o tym, że szybka reakcja oraz duża dokładność stanowiły o większych możliwościach wykonania znacznego obrotu w wyskoku. Zdolność wykonania obrotu w wyskoku w podobnym stopniu zależała od globalnych i lokalnych kondycyjnych zdolności motorycznych. Natomiast zdolności koordynacyjne były szczególnie istotne w swoich globalnych przejawach. Na tej podstawie można sądzić, że wyniki sprawdzianów stosowanych do kompleksowej oceny globalnych koordynacyjnych zdolności motorycznych zależą jedynie od poziomu niektórych zdolności motorycznych, mierzonych w zadaniach ruchowych o charakterze globalnym. W przypadku badanych studentów wartość obrotu w wyskoku, traktowana jako syntetyczny miernik koordynacji, zależała tylko od zdolności do szybkiej reakcji i zachowania równowagi oraz od zdolności różnicowania kinestetycznego. Natomiast nie było podstaw do wnioskowania we wpływie pozostałych zdolności koordynacyjnych na wynik tego testu.
Na podstawie analizy efektów badań własnych nadal nie można jednoznacznie określić przydatności pojedynczych zadań testowych do oceny koordynacyjnych zdolności motorycznych. Z jednej strony twierdzi się, że zastosowanie jednego testu do ich oceny nie dostarcza jednoznacznych wyników [Blume 1981; Raczek 1991a]. Z drugiej, istnieje szereg rezultatów badań, na podstawie których można wnioskować o istnieniu takich możliwości [Starosta 1978a]. Wydaje się jednak, że stosowanie jednego testu do oceny wszystkich zdolności koordynacyjnych może mieć nieco ograniczoną przydatność. Zdolności koordynacyjne zależą od wrodzonych możliwości przewodzenia impulsów nerwowych oraz od zasobu programów ruchu. Zdolności koordynacyjne przypuszczalnie należy badać w specyficznych zadaniach testowych – dostosowanych do wymagań koordynacyjnych, typowych dla wykonywanego zawodu lub uprawianej dyscypliny sportowej.
Koordynację ruchową można uznać za zdolność wybitnie specyficzną – związaną z ukształtowaniem określonych ośrodków i mechanizmów sterowania czynnościami ruchowymi. Natomiast u człowieka te ośrodki mogą być różne. Ponadto w trakcie procesu uczenia się określonych czynności ruchowych główne ośrodki ulegają zmianom. W procesie automatyzacji nawyku ruchowego zmniejsza się rola ośrodków korowych.
Także zdolność zachowania równowagi statycznej wyodrębniono do oddzielnej analizy. Powodem była jej szczególna rola w życiu codziennym człowieka i oczywiście w wychowaniu fizycznym i sporcie [Figura i wsp. 1991; Golema 1981, 1987; Wyżnikiewicz-Kopp 1992]. Badań nad zachowaniem równowagi prowadzono stosunkowo dużo, niestety nieliczne były opracowania, w których dążono do określenia związków zdolności zachowania równowagi z innymi przejawami zdolności motorycznych [Szeklicki i wsp. 1995]. Założono, że równowaga jest utrzymywana w ciągłym, dynamicznym procesie [Golema 1981; Hirtz 1985].
W badaniach własnych zdolności zachowania równowagi przy wykorzystaniu posturografu otrzymano wiele wskaźników wybitnie silnie wzajemnie powiązanych. Dlatego, za parametr wystarczająco reprezentatywny do jej oceny przyjęto pole powierzchni rozwiniętej przez rzut pionowy OSC. Stwierdzono, że równowaga mierzona na posturografie w standardowy sposób w niewielkim stopniu zależała od pozostałych zdolności motorycznych. Ze zdolności kondycyjnych warunkowały ją: czas narastania siły globalnej, prędkość lokomocyjna i częstotliwość globalna. Podobne rezultaty uzyskali Szeklicki, Osiński, Biernacki, Rauk, Kowalczyk, Kusy, Maciaszek [1995], gdzie równowaga była związana jedynie z rozmaitymi przejawami zdolności szybkościowych – lokalnych i globalnych. Natomiast w prezentowanych badaniach własnych lokalne zdolności kondycyjne nie miały żadnego wpływu na możliwość zachowania równowagi. Zdolności koordynacyjne miały nieco większe znaczenie. Było to zgodne z wynikami innych autorów [Szeklicki i wsp. 1995]. Różnice w znaczeniu ich globalnych i lokalnych przejawów były niewielkie. Ze zdolności koordynacyjnych największe znaczenie miała wielkość maksymalnego obrotu. Ujemny charakter zależności pomiędzy tą zdolnością a możliwością zachowania równowagi dowodził istnienia pomiędzy nimi pozytywnego związku. Mniejsza wartość pola powierzchni rozwiniętej przez rzut pionowy OSC była powiązana z możliwością wykonania większego obrotu w wyskoku.
Zdolność zachowania równowagi u człowieka odbywa się na drodze odruchowej [Golema, Jaśkiewicz 1986]. Natomiast jej poziom zależy głównie od właściwego współdziałania wzroku, zmysłu czucia głębokiego (zdolność różnicowania ruchu) i aparatu przedsionkowego ucha środkowego. Ponadto, istotne jest sprawne działanie układu nerwowo-mięśniowego [Golema 1987; Wyżnikiewicz-Kopp 1992]. Wydaje się, że czas narastania siły, który znacząco korelował ze zdolnością zachowania równowagi był uwarunkowany sprawnością funkcjonowania mięśni oraz zdolnością przewodzenia bodźców przez układ nerwowy. Ten ostatni element, podobnie jak i zdolność zachowania równowagi, wydaje się być niezależny od płci [Szopa 1992]. Ciało człowieka podczas utrzymywania równowagi traktowane jest jako obiekt regulacji, którym kieruje regulator, tj. układ nerwowy. Większa sprawność mechanizmu sterowania zachowaniem równowagi zależy od szybkości działania sprzężenia zwrotnego w układzie sterowania. Główną rolę w tej czynności odgrywa analizator kinestetyczny. Kierowanie ruchem przez układ nerwowy jest realizowane na trzech poziomach [Rutkowska-Kucharska i wsp. 1990]. W zadaniach złożonych, do których należy zachowanie równowagi, działanie odbywa się na podstawie programów wypracowanych przez wyższe piętra układu nerwowego. Jest ono powiązane z możliwościami wykonywania ruchów (np. manipulacyjnych), w których program budowany jest na bieżąco w trakcie ruchu – na zasadzie „on line”. Do podobnej interpretacji skłaniali się też Szeklicki, Osiński, Biernacki, Rauk, Kowalczyk, Kusy, Maciaszek [1995].
Prawdopodobnie uwarunkowań zdolności do zachowania równowagi w pozycji stojąc można poszukiwać w innych zdolnościach koordynacyjnych, zwłaszcza o globalnym charakterze aktywności ruchowej. Wydaje się, że większość zadań ruchowych o złożonej strukturze wykazuje powiązania z elementarnymi przejawami poszczególnych koordynacyjnych zdolności motorycznych. Natomiast te ostatnie wydają się być względnie niezależne [Ljach 1990, 2001; Mynarski 2000; Raczek i wsp. 2002]. W badaniach młodych piłkarzy nożnych Ljach i wsp. [2000] stwierdzili, że na 1518 współczynników korelacji aż w 85–95% nie było związków pomiędzy koordynacyjnymi zdolnościami motorycznymi. Pozostałe 5 do 15% istotnych zależności dotyczyło wskaźników w obrębie tych samych koordynacyjnych zdolności motorycznych.
Wydaje się, że przedstawione dane sugerują zjawisko wykorzystywania „małych programów ruchowych” (wzorców ruchów), związanych z umiejętnościami wykonywania określonych czynności ruchowych, do budowy „dużych programów ruchowych”, niezbędnych w realizacji kompleksów ruchowych. Przypuszczalnie proces ten przebiega w jedną, przedstawioną powyżej, stronę. Z tego powodu występują związki pomiędzy elementarnymi przejawami zdolności koordynacyjnych a zadaniami ruchowymi o złożonej strukturze, pomimo nielicznych zależności pomiędzy poszczególnymi podstawowymi przejawami zdolności koordynacyjnych.
Analizę stosowanych w badaniach własnych zdolności motorycznych wykorzystano do realizacji drugiego celu, tj. określenia związków występujących pomiędzy kondycyjnymi i koordynacyjnymi zdolnościami motorycznymi. W jej efekcie nie znaleziono podstaw do wnioskowania o istnieniu silnych zależności w tym zakresie. Zatem, pomimo poglądów o jedności procesów energetycznych i regulacyjnych, nie można wnioskować o istnieniu prostych powiązań pomiędzy ich przejawami [Krüger 1989; Vilkner 1989]. Stanowi to potwierdzenie drugiej hipotezy – o braku silnych koordynacyjnych uwarunkowań kondycyjnych zdolności motorycznych. Wydaje się, że na tej podstawie można zalecić odpowiedni dobór ćwiczeń do rozwijania obydwu grup zdolności motorycznych. W tym celu należy wykorzystywać zadania ruchowe o strukturze wymagającej zaangażowania procesów informacyjnych i energetycznych [Raczek i wsp. 2002]. Do podobnych wniosków doszła też Wyżnikiewicz-Kopp [1992], która w badaniach eksperymentalnych udowodniła, że równoczesne oddziaływanie na koordynacyjne zdolności motoryczne oraz zdolności siłowe i szybkościowo-siłowe znacząco wpłynęło na liczbę korelacji występujących pomiędzy nimi. Dlatego równoczesne rozwijanie potencjału kondycyjnych i koordynacyjnych zdolności motorycznych wpływa korzystnie na ich przejawianie w czynnościach sportowo-technicznych [Raczek 1990]. Należy przy tym uwzględnić większą specyficzność koordynacyjnych zdolności motorycznych. W praktyce wymaga to z jednej strony bardziej ukierunkowanego postępowania przy doborze środków treningowych. Z drugiej natomiast, zastosowania większej liczby rozmaitych ćwiczeń w procesie treningowym.
Zdolności motoryczne dzieli się najczęściej na kondycyjne i koordynacyjne, opierając się na koordynacyjnej złożoności zadania. Pomimo szeregu koncepcji, w których dążono do wprowadzenia obiektywnego podziału, ten rodzaj klasyfikacji ma charakter subiektywny. Nie uwzględnia się w nim zmian względnego odczucia stopnia komplikacji zadania, który jest zmienny i uzależniony od opanowania techniki ćwiczenia. Na przykład złożony element gimnastyczny dla nie uprawiającego tej dyscypliny sportu jest trudnym, lub niemożliwym do wykonania zadaniem, natomiast dla gimnastyka reprezentującego wysoki poziom sportowy stanowi łatwe lub średnio trudne ćwiczenie. Skomplikowane jest też oddzielenie zdolności kondycyjnych od koordynacyjnych. Przykładem tych problemów jest wyodrębnienie zdolności hybrydowych, tj. pośrednich pomiędzy kondycyjnymi a koordynacyjnymi [Raczek 1991a; Szopa 1989a, 1993; Mynarski 2000]. Z tych powodów wprowadzono ponownie pojęcie zwinności dla określenia czynności ruchowych, w których konieczne było wykazanie szybkości (podłoże energetyczne) oraz precyzji ruchów (podstawy koordynacyjne) [Czajkowski 1993; Raczek 1993]. Na podstawie wyników badań uznaje się za testy zwinności te zadania motoryczne, które w większym stopniu zależą od podłoża energetycznego niż informacyjnego [Szopa 1988; Raczek, Mynarski 1991, 1992; Szopa, Wątroba 1992].
Dlatego zaproponowano podział na zadania optymalne i maksymalne. Jego podstawą był cel działania. Nadrzędne stawało się uzyskanie określonego efektu. W zadaniach maksymalnych dążono do osiągnięcia jak najlepszego (maksymalnego) wyniku. Natomiast celem zadań optymalizacyjnych było przejawienie dokładnie określonej wartości. W pierwszym przypadku ocenie podlegała największa wartość wyniku a w drugim miarą była wielkość odchylenia od wartości zadanej – mniejszej od maksymalnej. Należy podkreślić, że w życiu codziennym i w sporcie często występują też sytuacje pośrednie, gdzie niezbędne jest uzyskanie maksymalnej efektywności działania przy pełnym wykorzystaniu możliwości energetycznych [Ozierov 1983]. W proponowanym podziale nie ma żadnego znaczenia stopień złożoności ruchu, warunkujący relacje pomiędzy potencjałem energetycznym a możliwościami układu nerwowego, dotychczas traktowany jako podstawowe kryterium podziału ćwiczeń na kondycyjne i koordynacyjne.
Podczas analizy wyników badań własnych stwierdzono, że precyzyjne przejawianie ruchów w pewnym stopniu zależało od maksymalnych wartości. Większość współczynników korelacji oraz regresji miało ujemny kierunek. Oznaczało to, że na ogół można spodziewać się, iż znaczne możliwości motoryczne – określane na podstawie maksymalnych rezultatów – determinowały znaczną zdolność precyzyjnego ich dawkowania, a więc dokładnego wykonywania ćwiczeń. Większość zadań stosowanych w niniejszych badaniach do oceny zdolności koordynacyjnych miała charakter optymalny. Dlatego zależności pomiędzy optymalnymi a maksymalnymi przejawami zdolności motorycznych pokrywały się ze związkami występującymi pomiędzy kondycyjnymi a koordynacyjnymi zdolnościami motorycznymi.
Związki pomiędzy zdolnością do osiągania maksymalnego wyniku a umiejętnością precyzyjnego przejawiania określonej wartości zdolności motorycznych najbardziej wyraźnie uwidoczniły się na przykładzie siły. Rozpatrując wartości maksymalnej siły globalnej i lokalnej, stwierdzono bardzo wyraźne różnice pomiędzy nimi. Prawdopodobnie, zarówno indywidualne wartości siły jak też rezultaty typowe dla każdego zadania motorycznego zależały od takich czynników, jak ilość włączonych do pracy jednostek motorycznych, przekrój fizjologiczny pracujących mięśni, rodzaj i wielkość włókien mięśniowych oraz wartość siły rozwijanej przez każdy mostek poprzeczny [Jaskólska 1998]. Istotne dla rozwinięcia znacznej siły jest też unerwienie jednostek motorycznych oraz możliwości wyzwolenia energii z fosfokreatyny [Szopa 1993; Popinigis 1997]. Zatem, dążąc do uzyskania siły maksymalnej, należy pobudzić jak największą liczbę jednostek motorycznych, stosując maksymalną częstotliwość bodźców [Jaskólska 1998]. Ważne jest też zsynchronizowanie aktywności jednostek motorycznych [Zaciorski 1970]. Ta umiejętność stwarza znaczne rezerwy w zakresie zwiększenia napięcia mięśniowego. U nie trenujących w sposób synchroniczny działa od 20% do 50% (w małych mięśniach) impulsów. Natomiast poprzez odpowiedni trening można tę wielkość znacznie powiększyć [Zaciorski 1970].
Różnice w sile typowej dla zadań o globalnym i lokalnym charakterze były oczekiwane. Natomiast bardzo interesujące wydaje się podobieństwo czasów osiągania maksymalnych wartości globalnej siły statycznej i lokalnej siły statycznej. Oznacza to, że zdolność do szybkiego narastania siły może być wartością indywidualną, uzależnioną od predyspozycji osobniczych. Nie ma na nią wpływu rodzaj ćwiczenia oraz liczba zaangażowanych w nim grup mięśniowych. Prawdopodobnie determinują ją proporcje występujące pomiędzy szybkimi i wolnymi włóknami mięśniowymi [Celichowski 1995]. Wydaje się, że zależy ona od kinetyki mostków poprzecznych [Jaskólska 1998]. Szybkość kurczenia się włókien mięśniowych jest uwarunkowana szybkością przechodzenia mostka w stan rozwijania siły [Huxley, Simmons 1972]. Szczególne znaczenie ma szybkość dochodzenia jonów wapnia do cienkich filamentów [Jaskólska 1998]. Ponadto zdolność do szybkiego narastania siły jest uzależniona od rekrutacji jednostek motorycznych i sztywności elementów sprężystych. Dużą rolę odgrywa także liczba mostków położonych równolegle względem siebie i równocześnie aktywowanych [Mero i wsp. 1981; Szczęsna i wsp. 1996; Viitasalo, Komi 1981]. Należy podkreślić też znaczenie zmian częstotliwości z wzorca wyładowań motoneuronów czynnych jednostek [Celichowski 1995]. Ten mechanizm ma szczególne znaczenie podczas wykonywania precyzyjnych ruchów. Natomiast w zadaniach o ogólnym charakterze najważniejsze wydaje się być pobudzenie odpowiedniej liczby jednostek [Celichowski 1995].
Stwierdzono w badaniach własnych, że w zadaniach wymagających optymalnego przejawienia zdolności motorycznych, w których określano dokładność siłową, badani osiągali czasy jedynie niewiele krótsze od typowych dla siły maksymalnej. Zatem uzyskanie zadanej siły było zadaniem bardziej złożonym niż przejawienie jej maksymalnej wartości. Przypuszczalnie wiąże się to z koniecznością stworzenia przez badanych odpowiedniego programu działania i dłuższym czasem jego realizacji [Starosta i wsp. 1994; Rynkiewicz, Starosta 2000]. W zadaniach wymagających dokładności siłowej na początku zadania siła przyrastała stosunkowo powoli [Rynkiewicz, Starosta 1999, 2000]. Zatem odwrotnie niż w pomiarach jej największej wartości, gdzie początek wzrostu był najszybszy [Clarke 1968]. Prawdopodobnie podstawą wykonania precyzyjnego ruchu było jego odpowiednie zaprogramowanie, a dopiero w następnej kolejności realizacja wymagająca zaangażowania mechanizmów energetycznych [Pöhlmann 1986]. To zjawisko było podobne w pomiarze globalnej i lokalnej dokładności siłowej. Szybkość narastania siły mniejszej od połowy wartości maksymalnej była uzależniona od rekrutacji jednostek motorycznych [Milner-Brown, Stein 1973]. Wydaje się zatem, że programowanie wykonania zadania wymagającego dokładności siłowej polega głównie na wyborze odpowiedniej liczby jednostek motorycznych oraz właściwej dla danego osobnika częstotliwości bodźców. Zaciorski [1970] twierdził ponadto, że dozowanie napięcia mięśniowego w wysiłkach mniejszych od maksymalnych przebiega inaczej niż w zadaniach o maksymalnym oddziaływaniu. Wyższe ośrodki nerwowe odpowiadają za określenie przestrzennych, czasowych i szybkościowych wielkości charakteru ruchu. Natomiast wybór odpowiednich kombinacji napięć mięśniowych leży w gestii niższych ośrodków. Ponadto, impulsy nerwowe wysyłane są najpierw do części ruchowej wrzecion mięśniowych. Efektem tego jest zmiana ich napięcia i odpowiednie pobudzenie części czuciowej. W dalszej części regulacja odbywa się zgodnie ze schematem odruchu na rozciąganie. Natomiast w zadaniach wymagających maksymalnego napięcia mięśniowego część ruchowa wrzecion mięśniowych, unerwiona przez system gamma, nie ma znaczenia, ponieważ bodźce wysyłane są z ośrodków mózgu poprzez komórki ruchowe rdzenia bezpośrednio do włókien mięśniowych. W obydwu przypadkach regulatorem są ośrodki korowe półkul mózgowych.
Prawdopodobnie odpowiedni trening może być czynnikiem skracającym czas narastania siły w ćwiczeniach optymalnych. Wartość tego czasu powinna dążyć do wielkości typowych dla czasu przejawiania siły maksymalnej. Jego skracanie się winno następować poprzez uczenie się szybkiego i efektywnego konstruowania programów dokładnego wykonania ruchów. Wymaga to wzbogacenia doświadczeń (zwiększenia zasobu programów wykonywania ruchów) poprzez stosowanie odpowiednich ćwiczeń, które powinny być nowe albo zawierać elementy nowości [Zaciorski 1970; Ulatowski 1981; Hirtz 1985].
W zależności od rodzaju zadania motorycznego zmienia się umiejętność precyzyjnego dozowania jego intensywności, czyli możliwość przejawiania w sposób optymalny poszczególnych zdolności motorycznych. Dokładność ruchów jest podstawowym kryterium oceny poziomu zdolności koordynacyjnych [Raczek 1991a; Górgól 1993]. Pomimo tego stwierdzenia analiza dokładności ruchów w praktyce była rzadko prowadzona, chociaż znaczenie umiejętności precyzyjnego wykonywania ruchów było i pozostaje bardzo duże, a jej rola była podkreślana przez wielu autorów [Grottel, Celichowski 1992]. Dokładność ruchów zależy od zdolności różnicowania kinestetycznego, która jest uznawana za jedną z podstawowych zdolności motorycznych [Blume 1981; Harre 1985; Hotz, Weineck 1983; Hirtz 1985; Ljach 1990, 2001; Raczek, Mynarski 1992; Szopa 1993; Starosta 1993a, 2001a; Szopa i wsp. 1996; Mynarski 2000; Raczek i wsp. 2002]. Jest ona w dużym stopniu uwarunkowana genetycznie. Chociaż istnieją pewne możliwości jej poprawienia poprzez odpowiedni trening [Szopa 1993; Szopa i wsp. 1996; Prus 2000]. Przejawienie dokładności zależy od umiejętności sterowania czynnością poszczególnych mięśni i grup mięśniowych. Te z kolei są uwarunkowane możliwościami przyjmowania i przetwarzania informacji o kątowej pozycji w stawach (składowe przestrzenne) oraz o napięciu pracujących mięśni (składowe siłowe) i prędkości ruchów (składowe czasowe) [Mynarski 2000]. Warunkują ją też możliwości rekrutacji jednostek motorycznych w skurczu oraz częstotliwość pobudzania włókien nerwowych i działanie mięśni antagonistycznych.
Ze względu na praktyczne znaczenie osiągania odpowiedniej dokładności ruchów, w badaniach własnych porównano jej przejawianie się w licznych zadaniach motorycznych. Uzyskane wyniki uporządkowano zgodnie z wartościami typowego dla nich błędu przejawiania zadanej wartości. Stwierdzono ich duże zróżnicowanie. Kierując się wartością błędu, wszystkie zadania motoryczne podzielono na cztery grupy. W pierwszej, o błędach mniejszych niż 10%, znalazły się zadania wymagające dokładnego przejawienia siły w pojedynczych ruchach lokalnych i globalnych. Do drugiej grupy zakwalifikowano te, w których konieczne było precyzyjne odtworzenie częstotliwości ruchów. Natomiast do trzeciej grupy zaliczono zadania ruchowe polegające na wykonywaniu pojedynczych ruchów o lokalnym lub globalnym charakterze aktywności mięśniowej. Najmniej dokładnie odtwarzano 50% wartości maksymalnej prędkości w biegu na odcinku 5 m (ryc. 11). To ostatnie zadanie zaliczono do ostatniej, czwartej grupy.
Największą dokładność uzyskano w zadaniach, w których układano program ruchu przed jego rozpoczęciem, natomiast w trakcie realizacji nie było możliwości dokonania znaczącej korekty. Były to zadania wymagające przejawienia dokładności odtworzenia określonej wartości obrotu w wyskoku oraz osiągnięcia zadanej wysokości wyskoku pionowego. Czas ich wykonania nie przekraczał 0,2 s. W tego rodzaju ruchach nie ma znaczących możliwości korygowania ich programu w trakcie realizacji – przynajmniej w jego początkowej fazie [Bernstein 1975; Kulig i wsp. 1978]. Zatem dokładność w ruchach balistycznych zależała głównie od poprawności programu wykonania zadania, skonstruowanego przed jego rozpoczęciem. Natomiast duża dokładność przejawienia globalnej i lokalnej maksymalnej siły statycznej mogła być efektem systematycznego wprowadzania korekt do zadań trwających około 0,6–0,8 s. Było to tym prostsze, że decydujące o wyniku przyrosty siły zachodziły w końcowej fazie wykonania zadania ruchowego [Rynkiewicz, Starosta 2000]. Czynnikiem ułatwiającym było też odbieranie bodźców tylko z jednego rodzaju receptorów, które w pomiarze siły lokalnej pochodziły głównie z jednej grupy mięśniowej – zginaczy stawu łokciowego. Prawdopodobnie dlatego najbardziej dokładnie odtwarzano wartość maksymalnej lokalnej siły statycznej.
Mniej dokładne wyniki przejawiono w zadaniach ruchowych typowych dla drugiej grupy – wykonywanych z wysoką częstotliwością. Przypuszczalnie znaczna częstotliwość ruchu wpłynęła na ograniczenie korygowania jego programu z powodu napięcia szybkościowego mięśni, które na ogół towarzyszy ruchom szybko po sobie następującym.
Zadania ruchowe charakterystyczne dla trzeciej grupy zawierały pojedyncze ruchy o lokalnym i globalnym charakterze aktywności mięśniowej. Wyniki ich wykonania odznaczały się stosunkowo małą dokładnością odtworzenia zadanej intensywności. Czas wykonania tych zadań był podobny jak w pierwszej grupie, a jednak uzyskano gorsze rezultaty. Prawdopodobnie przyczyn należy upatrywać w dynamicznym charakterze aktywności mięśniowej. W pierwszej grupie zadania realizowano w warunkach statyki lub quasi-statyki – odbicie w wyskoku.
Wydaje się, że prawidłowe korygowanie programu wykonania ruchu, które umożliwia włączenie analizatora wzrokowego, nie wpływało korzystnie na dokładność ruchu. Najniższe rezultaty były typowe dla zadania cyklicznego, w którym poza kontrolą wzrokową można było też korygować intensywność ruchu dzięki analizie przemieszczania się poszczególnych części ciała – prędkość na odcinku 5 m.
Według Bernsteina czas obiegu informacji w organizmie wynosi od 0,07 s do 0,12 s. Oznacza to częstotliwość rzędu 8–14 Hz, która jest typowa dla rytmu α fal mózgowych i tremoru mięśniowego. Na podstawie analizy szybkości obiegu informacji od receptora do efektora można twierdzić, że ruchy balistyczne są sterowane „ante factum”. Natomiast w ruchach ciągłych sterowanie odbywa się na zasadzie „in facto” [Morecki i wsp. 1971]. Sterowanie ruchem odbywa się na zasadzie nadążnego porównywania SW (Sollwert – wyobrażenie ruchu) z wartością IW (Istwert – realna wartość przebiegu ruchowego). Pobudzanie mięśni wymaga stałego dopasowywania się ΔW (wartości różnicowe) do zaistniałej sytuacji, tak aby możliwe było wykonanie zadania ruchowego. Oczywiście konieczna jest wola do realizacji ruchu lub do jego przerwania w przypadku niemożności jego wykonania [Grottel, Krutki 1996].
Dokładne i ekonomiczne wykonanie ruchu było uwarunkowane precyzją i obiektywnym odbiorem, prawidłową oceną oraz przetwarzaniem informacji o siłowych, przestrzennych i czasowych wskaźnikach ruchu. Dla przejawienia najwyższej dokładności ruchu konieczne jest posiadanie informacji o stanie napięcia poszczególnych grup mięśniowych, prędkości ruchów oraz pozycji kątowej w stawach. Precyzyjne „czucie” tych elementów ruchu umożliwia właściwe dozowanie impulsów sterujących, niezbędne dla uzyskania optymalnego przebiegu ruchu.
Według Bernsteina czynności ruchowe są samoregulującymi się procesami, które mogą funkcjonować tylko przy stałym wykorzystywaniu informacji na zasadzie sprzężenia zwrotnego. W realizacji ruchów duże znaczenie ma samokontrola i kierowanie ruchem przez ćwiczącego. Planowanie i realizacja dowolnego, zamierzonego ruchu jest możliwa jedynie dzięki stałemu śledzeniu i korygowaniu jego przebiegu za pomocą organów czuciowych [Bernstein 1966]. Wykonanie zadań ruchowych wymaga uwzględnienia czynników zewnętrznych i wewnętrznych oraz zastosowania informacji o nich na zasadzie „aferentacji zwrotnej” oraz „korekcji sensorycznej” [Bernstein 1966; Ozierov 1983]. Zdolność do kierowania ruchem i samokontroli ćwiczącego jest ważna i uwzględniana w sporcie, psychologii pracy i przygotowaniu zawodowym [Czebyszeva 1969; Kossov 1973, 1976].
Podstawową funkcją samokontroli człowieka jest śledzenie przez jego system kierujący stopnia zgodności wykonywanego ruchu z jego programem [Kossov 1976; Ozierov 1983]. Zgodnie z poglądami Łomova, Kossova i Konopkina [1975, za Ozierovem, 1983] samokontrola w zakresie kierowania ruchem powinna spełniać następujące założenia:
Ćwiczący powinien umieć różnicować poszczególne parametry ruchu, ponieważ o precyzji ruchu stanowią możliwości określania małych rozbieżności pomiędzy programem a jego realizacją.
Ćwiczący powinien dysponować zdolnością do szybkiego przesyłania informacji niezbędnych do kierowania ruchem, co z kolei jest uwarunkowane możliwościami szybkościowymi układu nerwowego.
Ćwiczący powinien umieć ukierunkować samokontrolę na określone fazy i elementy ruchu.
Struktura samokontroli kierowania ruchem powinna zależeć od sposobu analizy elementów ruchu (świadomie lub automatycznie), podstawy sensorycznej (kinestetyczna, wzrokowa i in.) oraz od zmienności ruchu (przestrzenna, dynamiczna, czasowa).
Samokontrola musi być stabilna, czyli możliwa do realizacji w warunkach zakłóceń wewnętrznych lub zewnętrznych (emocje, zmęczenie, czynniki zewnętrzne).
Wszystkie ćwiczenia powinny być przedmiotem stałej samokontroli sposobu ich wykonywania. Natomiast wybór elementów wykorzystywanych do niej zależy od rodzaju ruchu oraz od indywidualnych predyspozycji ćwiczących. Ich dobór ma wpływ na czas niezbędny do nauki danego zadania ruchowego. Samokontrola może być zautomatyzowana lub świadoma. Jej rodzaj zależy także od rodzaju ruchu i stopnia jego opanowania [Ozierov 1983].
Wyższy poziom czucia sensorycznego charakteryzuje sportowców, a niższy osoby nie trenujące [Farfel 1960; Puni 1963, 1967]. Czucie ruchowe (sensoryczne) zmienia się u sportowców proporcjonalnie do wzrostu poziomu ich formy sportowej oraz mistrzostwa technicznego [Ozierov 1983; Anioł-Strzyżewska, Starosta 1984]. Ponadto, przejawianie precyzyjnego czucia ruchu zależy od rodzaju uprawianej dyscypliny sportowej [Ozierov 1983; Starosta 1983].
W rozwoju nawyku ruchowego podstawową rolę odgrywa kształtowanie rytmu i tempa ruchów, które determinują możliwość wykonania czasowo-przestrzennej struktury ruchu [Ozolin 1970; Zaciorski 1970; Djaczkov 1975]. Opanowanie tempa i rytmu jest niemożliwe bez odpowiedniego czucia czasu, czyli umiejętności precyzyjnego rozróżniania parametrów czasowych oraz różnicowania swoich ruchów w dokładnie zadanym czasie [Gellersztejn 1958]. W sporcie bez tej umiejętności trudno jest odnieść sukces [Rudik 1958]. Czucie czasu odbywa się dzięki funkcjonowaniu różnych receptorów, głównie słuchowego i kinestetycznego [Ozierov 1983, za Sjeczenovem 1947]. Jest uzależnione od wzajemnych powiązań procesów pobudzenia i hamowania na poziomie korowym [Ozierov 1983 za Woodworth 1963; Krech, Crutchbled, Livson 1969]. Może się zmieniać w zależności od stanu emocjonalnego ćwiczących oraz od zaangażowania energetycznego, np. od siły niezbędnej do wykonania ruchu a także od poziomu zaawansowania sportowego zawodnika [Gellersztejn 1958].
Dokładność czasowa jest bezpośrednio związana z dokładnością przestrzenną i siłową. Dokładność przestrzenna jest konieczna do opanowania techniki ruchu [Puni 1967; Djaczkov 1975; Ozolin 1970]. Zastosowanie natychmiastowej informacji o wykonywanych ruchach umożliwia znaczące poprawienie oceny przestrzennych i czasowych parametrów ruchu, a tym samym szybsze jego opanowania [Farfel 1960; Czchaidze 1970; Donskoj, Zaciorski 1979].
Nie dotyczy to zadań ruchowych o czasie realizacji krótszym niż 0,12 s – ruchy balistyczne. W tych przypadkach ruch odbywa się bez możliwości automatycznej korekty i uwarunkowany jest tylko biomechanicznymi właściwościami organizmu ćwiczącego [Agaszin 1977]. Sterowanie nim odbywa się na zasadzie „ante factum” [Morecki i wsp. 1971]. W pozostałych przypadkach mamy do czynienia z „psychomechaniczną” sferą działalności człowieka [Agaszin 1977]. Do sterowania ruchem wymagana jest odpowiednia struktura informacyjna, rozumiana jako przekazywanie szeregu uporządkowanych w czasie danych, zawierających ustaloną ilość informacji o ruchu i warunkach jego realizacji. Każda dynamiczna, kinematyczna lub rytmiczna struktura ruchu zawiera część informacyjną. Można twierdzić, że przekazanie informacji o parametrach ruchu na poszczególne części ciała człowieka w określonym momencie stanowi przestrzenną część struktury informacyjnej, a zmiany tych informacji w trakcie ruchu stanowią jej składową czasową. Istnienie tej struktury pozwala na wnioskowanie o „psychomechanicznej” strukturze ruchów człowieka [Agaszin 1977].
Dokładność siłowa stanowi ważną zdolność warunkującą prawidłowe nauczanie ruchów [Puni 1957]. Stwierdzono, że poprawia się ona zdecydowanie szybciej niż dokładność czasowa i przestrzenna [Mieńszikov 1966]. Natomiast brakuje opracowań dotyczących zależności pomiędzy różnymi rodzajami dokładności [Leonov 1973]. W badaniach własnych stwierdzono istnienie stosunkowo nielicznych powiązań pomiędzy niektórymi globalnymi zdolnościami motorycznymi w zadaniach wymagających przejawienia dokładności. Natomiast w obrębie optymalizacyjnych zdolności motorycznych o charakterze lokalnym zaobserwowano tylko jeden statystycznie znaczący współczynnik korelacji. Stanowiło to kolejne potwierdzenie założenia, że koordynacyjne zdolności motoryczne, a szczególnie ich lokalne przejawy, są względnie niezależne.
Można sądzić, że realizując trzeci cel, którym było określenie powiązań pomiędzy optymalnym (dokładnym) a maksymalnym przejawianiem się zdolności motorycznych, uzyskano informacje o istnieniu uwarunkowań możliwości dokładnego wykonywania ruchu maksymalnymi możliwościami motorycznymi. Stwierdzono też, że poszczególne zadania motoryczne charakteryzowały się zróżnicowaną dokładnością.Potwierdzono tym samym trzecią hipotezę. Sądzimy, że to zjawisko można wytłumaczyć, korzystając z opracowanej przez Bernsteina [1947] teorii sterowania i regulacji czynności ruchowych. Zgodnie z podstawowymi założeniami wielopoziomowej teorii Bernsteina [1947], dla realizacji zadań ruchowych o rozmaitym celu i charakterze, wymagających różnych cech psychofizjologicznych, tworzy się wielopoziomowe konstrukcje. W każdej z nich zasadniczą funkcję regulacyjną pełni jeden poziom główny, na którym określa się racjonalne aspekty i cele aktywności ruchowej. Ten poziom odgrywa podstawową rolę w wykonaniu czynności ruchowej, w której uczestniczą pozostałe poziomy odpowiedzialne za drugorzędne składniki ruchu, takie jak napięcie mięśni czy ich współdziałanie. Każdy poziom ma własny składnik anatomiczny w określonym miejscu centralnego układu nerwowego oraz typowe dla siebie korekty czuciowe, będące podstawą koordynacji ruchów. Według Bernsteina [1947, 1966] zakres rozwoju poszczególnych poziomów koordynacyjnych jest indywidualnie zróżnicowany. Jednak stopień rozwoju i możliwości wytrenowania są swoiste, nie tylko dla poszczególnych aktów ruchowych, ale dla odpowiednich grup ruchów wykonywanych dzięki istnieniu właściwego poziomu konstrukcji w centralnym układzie nerwowym. Ponieważ liczba tychże jest niewielka, łatwiej jest je oceniać niż badać oddzielnie poszczególne akty ruchowe.
Ljach [1987, 1990] na podstawie badań ponad 2000 dzieci w wieku 7–17 lat, u których analizował około 5000 wskaźników korelacji, udowodnił, że dodatni transfer zdolności koordynacyjnych można stwierdzić tylko w czynnościach ruchowych, konstruowanych przy jednakowych poziomach głównych i drugorzędnych. Dodatni, lecz nieco mniejszy transfer odnotowano w przypadku podobnych poziomów wiodących i zróżnicowanych poziomów tła. Natomiast przy rozmaitych poziomach głównych i drugorzędnych nie ma żadnych zależności, a nawet pojawiają się związki negatywne [Ljach 1990, 1991a, 2001].
Ljach na podstawie przeprowadzonych badań uważał, że przy niewielkim, ale stosunkowo równym doświadczeniu ruchowym, przy niezbyt dużej liczbie zautomatyzowanych składowych ruchu, poziom dokładności wykonania zadań motorycznych zależy od stopnia rozwoju zasadniczych poziomów konstrukcji ruchu, określających racjonalne i programowe elementy ruchów. Poprawa doświadczenia ruchowego i automatyzacja coraz większej liczby składowych czynności ruchowych spowoduje, że wyższy stopień zdolności koordynacyjnych wykażą osoby o szerszym zakresie i wyższym stopniu opanowania nawyków technicznych oraz większej automatyzacji na tworzących tło poziomach pola przestrzennego C i poziomu synergii B. Ich łączne działanie określa szybkość i racjonalność – podstawowe kryteria oceny zdolności koordynacyjnych w końcowym etapie uczenia się złożonych koordynacyjnie czynności ruchowych [Ljach 1990, 1991a, 2001].
Symetryczność przejawiania zdolności motorycznych jest bardzo ważnym zagadnieniem, chociaż nie do końca zbadanym [Starosta 1975, 1990, 1993b, 2001a; Drozdowski S. 1980; Drabik 1982b, 1983; Raczek i wsp. 2002]. Uznawana jest za jeden z podstawowych problemów do rozwiązania w zakresie nauk o zdolnościach motorycznych [Starosta 1975, 1990, 2001a]. Jej rozwój jest bardzo istotnym zadaniem w kształtowaniu wysokiego poziomu techniki sportowej i koordynacji ruchowej. Lateralizacja lub stronne zróżnicowanie, rozumiane jako preferowanie jednej strony lub kończyny, jest wrodzone i uzależnione od asymetrycznych właściwości mózgu. W pewnym stopniu może też być efektem odpowiedniego treningu [Stokłosa 1998].
Niezależnie od indywidualnie wybranej kończyny dominującej, niezbędne jest obustronne wykonywanie czynności ruchowych. Stanowi to jedno z podstawowych uwarunkowań podniesienia poziomu koordynacyjnych zdolności motorycznych [Ljach 1990, 1995; Starosta 1975, 1990, 1995b; Zaciorski 1979; Raczek i wsp. 2002].
W dążeniu do realizacji kolejnego – czwartego – celu, którym było określenie stopnia symetryczności przejawiania ocenianych zdolności motorycznych, stwierdzono w badaniach własnych, że zdolności motoryczne rozwijane przy symetrycznym przejawianiu ruchów miały podobne wartości, szczególnie w zakresie zdolności koordynacyjnych. Statystycznie znaczące różnice pomiędzy wartościami średnich arytmetycznych odnotowano w czterech przypadkach. Dotyczyły one: prędkości pojedynczego ruchu, częstotliwości ruchów kończyną górną i dokładności w tym zakresie oraz maksymalnej siły lokalnej. Wszystkie one należały do zdolności kondycyjnych o charakterze lokalnym. Można przypuszczać, że jedynie lokalne kondycyjne zdolności motoryczne cechowała znaczna lateralizacja.
Natomiast prawie wszystkie wyniki zadań motorycznych, wykonywanych w jedną lub drugą stronę, prawą bądź lewą kończyną, były wzajemnie skorelowane. Wyjątek stanowiły dwie lokalne zdolności motoryczne, stanowiące o precyzji ruchów kończyny górnej. Były nimi lokalna dokładność siłowa oraz dokładność częstotliwości ruchów lokalnych. Przypuszczać należy, że różnice w przejawianiu się tych zdolności motorycznych obydwu kończyn górnych zostały spotęgowane ich rozmaitą rolą w wykonywaniu czynności życiowych, a zatem miały bardziej charakter wyuczony niż wrodzony. Pomimo znacznych podobieństw pomiędzy rezultatami typowymi dla każdej strony można twierdzić, że nieco bardziej korzystnie prezentowały się wyniki zadań wykonywanych w prawo. Pomimo tych uwag wydaje się, że potwierdzono czwartą hipotezę – o znacznych możliwościach symetrycznego przejawiania kondycyjnych i koordynacyjnych zdolności motorycznych, szczególnie o globalnym charakterze.
Wydaje się jednak, że globalny lub lokalny charakter zadania ruchowego nie wpływał znacząco na symetryczne przejawianie się zdolności motorycznych. Bez większego znaczenia była też dominacja uwarunkowań kondycyjnych lub koordynacyjnych oraz wymagań dotyczących maksymalnego lub optymalnego przejawiania aktywności (ryc. 17).
Rycina 17 Zestawienie wskaźników symetrycznego przejawiania zdolności moto-rycznych w zespole badanych studentów (n = 176)
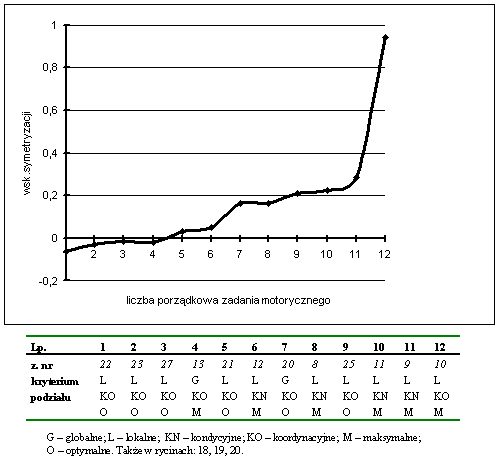
W przypadku lokalnych zdolności motorycznych określono wyraźną tendencję do bardziej symetrycznego przejawiania ich optymalnych przejawów, związanych z dokładnością wykonania zadania, w porównaniu z ich maksymalnymi wartościami (ryc. 18). I tak, dokładność wykonania pojedynczego ruchu i odtworzenia zadanej częstotliwości ruchów była bardziej symetryczna niż jego prędkość i częstotliwość maksymalna. Podobne zjawisko było też charakterystyczne dla lokalnej dokładności siłowej i lokalnej siły maksymalnej. Natomiast czasy osiągania siły lokalnej były zbliżone w pomiarze siły maksymalnej i dokładności siłowej.
Dla porównania zastosowano też obrót w wyskoku, czyli zadanie motoryczne wymagające globalnego przejawienia się koordynacyjnych zdolności motorycznych. Stwierdzono, że bardziej symetryczne wyniki uzyskano w wersji maksymalnej niż optymalnej (ryc. 18). Dowodzi to większej symetryczności w przejawianiu lokalnych zdolności motorycznych niż ich globalnych postaci, które wydają się być bardziej symetryczne wówczas, gdy wymagane jest osiągnięcie ich maksymalnej wartości.
Rycina 18 Zestawienie wskaźników symetryzacji w parach zadań motorycznych wymagających maksymalnej i optymalnej aktywności (n = 176)
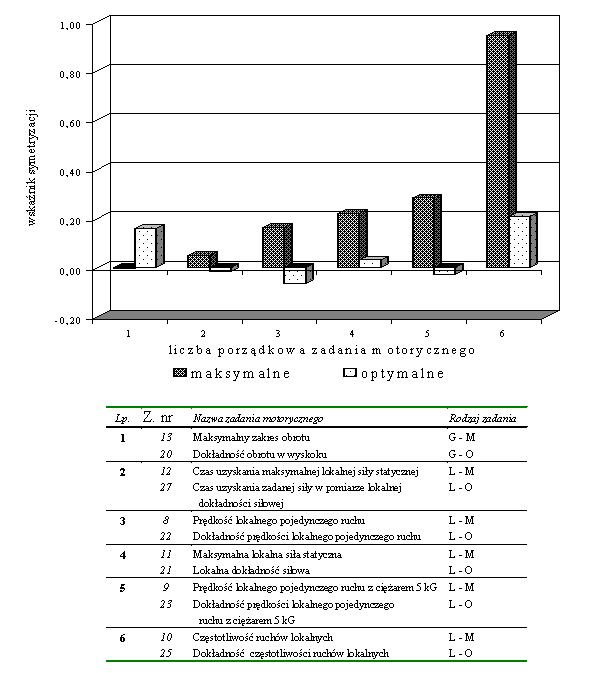
Wydaje się, że większe podobieństwo wyników typowych dla lokalnych zadań motorycznych wymagających dokładności niż odpowiadających im zadań o maksymalnej aktywności można uzasadnić dwoma powodami. Pierwszym, związanym z funkcjonowaniem układu nerwowego, wydaje się być mniejsza lateralizacja jego możliwości w zadaniach ruchowych wykonywanych bez ograniczenia czasowego (zadania optymalne) niż w warunkach dążenia do osiągnięcia najkrótszego czasu realizacji pojedynczego ruchu lub serii ruchów, typowych dla zadań o maksymalnym charakterze. Drugim powodem większej asymetrii w prostych ruchach mogą być ograniczenia w szybkości mobilizacji beztlenowych możliwości energetycznych na poziomie lokalnym.
Różnice w możliwości symetrycznego przejawiania zdolności motorycznych, typowe dla lokalnych zadań ruchowych o złożonej strukturze, można wyjaśnić, opierając się na wielopoziomowej teorii Bernsteina [1966, 1991]. Zgodnie z nią wszystkie ruchy wykonywane na pierwszych trzech poziomach są symetryczne. Natomiast czynności typowe dla czwartego poziomu, właściwego tylko człowiekowi i kształtującego się wraz z jego rozwojem, zawsze cechują się lepszymi wynikami prawej strony. Bernstein [1966, 1991] tłumaczy to dominacją u większości ludzi lewej półkuli mózgowej. Z badań własnych wynika jednak, że w większości zadań motorycznych typowy był znaczny stopień symetryzacji. Zwiększenia lateralizacji można oczekiwać w zadaniach ruchowych obciążających zarówno układ nerwowy, jak i mechanizmy energetyczne (maksymalna częstotliwość ruchów lokalnych) oraz w typowych działaniach z poziomu czwartego w klasyfikacji Bernsteina [1966, 1991].
Odpowiednio realizowany proces treningowy może spowodować zmniejszenie lateralizacji. Dowodzą też tego badania Stokłosy i innych autorów, na których ona się powołuje. Wynika z nich, że wśród sportowców zmniejsza się liczba osób o nieustalonej lateralizacji [Stokłosa 1998; Starosta 1975, 1990, 1993b, 1995b]. W procesie wychowania fizycznego i treningu sportowego należy poświęcać wiele uwagi symetrycznemu rozwijaniu zdolności motorycznych. Podstawą powinno być nauczenie wzorca ruchu w bardziej sprawną stronę. Następnie konieczne jest kształcenie umiejętności jego wykonania „słabszą” stroną lub kończyną. W tym celu należy stosować ćwiczenia polegające na tzw. „lustrzanym” wykonaniu [Zaciorski 1970; Starosta 1975, 1990; Raczek i wsp. 2002]. Kształtowanie symetrii ruchów jest szczególnie ważne w dyscyplinach, w których występuje bezpośredni kontakt z przeciwnikiem, a zatem w grach sportowych i w sportach walki [Zaciorski 1979; Starosta 1984, 1990, 1993b; Ljach 1990]. Zgodnie z poglądami Starosty [1984, 1990, 1993b] symetria ruchu wpływa korzystnie na koordynację i poziom mistrzostwa technicznego. Zdolność do symetrycznego przejawiania zdolności motorycznych jest pozytywnie związana z możliwościami opanowywania nowych nawyków ruchowych, niezbędnych do sprawnego wykonywania różnych zawodów, szczególnie związanych z operowaniem maszynami i urządzeniami.
Zróżnicowanie płciowe w zakresie zdolności motorycznych było przedmiotem licznych badań [Geblewiczowa, Ogórek 1960; Wolański, Pařizkowa 1976; Dannhauer 1977; Janusz, Jarosińska 1981; Pańczyk 1981; Drabik 1982b; Osiński, Swornowski 1983; Szopa i wsp. 1985; Arska-Kotlińska, Drozdowski Z. 1995; Pawłowa-Starosta, Starosta 1999]. Dymorfizm płciowy pojawia się w okresie płodowym i zwiększa się w trakcie rozwoju, osiągając maksymalną wartość u dorosłych. Natomiast w okresie starości zmniejsza się [Wolański 1983]. Chociaż istnieją też poglądy, poparte wynikami badań, że w zakresie cech psychomotorycznych kobiety mogą mieć wyższe rezultaty. Wiąże się to z okresem pokwitania [Szopa i wsp. 1985].
Większość opracowań dotyczyła zróżnicowania płciowego w zakresie kondycyjnych zdolności motorycznych. Badania nad przejawianiem się zwinności u kobiet i mężczyzn prowadził Osiński [1988b]. Natomiast Szopa, Mleczko i Cempla [1985] poza zdolnościami kondycyjnymi mierzyli też zdolności psychomotoryczne. Do oceny czasu reakcji zastosowali aparat Piórkowskiego, a do pomiaru orientacji przestrzennej aparat krzyżowy. Podobną procedurę zastosował też Mleczko [1991]. Nie stwierdzono natomiast opracowań traktujących o różnicach pomiędzy kobietami i mężczyznami w aspekcie podziału zdolności motorycznych na ich przejawy globalne i lokalne.
W badaniach własnych zaobserwowano wiele znaczących różnic w zakresie zdolności motorycznych występujących pomiędzy kobietami i mężczyznami. Do ich oceny wykorzystano analizę istotności różnic pomiędzy wynikami średnimi oraz wskaźnikami dymorfizmu.
Potwierdzono oczywiście zdecydowanie wyższe wyniki mężczyzn niż kobiet w zakresie zdolności kondycyjnych, zwłaszcza globalnych. Prawdopodobnie podłożem takiej tendencji była budowa ciała i jego skład determinujący zróżnicowanie tych zdolności. Istotne znaczenie mógł też mieć typ układu nerwowego [Yandell, Spirduso 1981]. Kobiety osiągały zdecydowanie niższe wyniki w zakresie siły maksymalnej i skoczności. Przypuszczalnie wpłynęła na to wielkość masy mięśniowej oraz układ enzymatyczny włókien i zawartość androgenów [Tanner 1963; Wilkie 1974; Wolański 1983]. Jaskólska, Adach, Nieścieruk-Szafrańska [1987] stwierdzili bardziej wyraźne różnice wyników kobiet i mężczyzn w zadaniach ruchowych wymagających większego udziału zdolności szybkościowo-siłowych. Ustalili też, że charakter wymagań względem motoryki stawiany przez dyscyplinę sportową warunkuje różnice między kobietami a mężczyznami. Ich wartość była proporcjonalna do wymagań szybkościowo-siłowych.
W badaniach własnych stwierdzono, że zdolności wymagające szybkiej aktywności układu nerwowego były podobne. Zaliczono do nich czas reakcji oraz czasy osiągania siły maksymalnej, zarówno lokalnej jak i globalnej. Także w zadaniach wymagających przejawiania optymalnych wartości stwierdzono nieliczne różnice. Kobiety cechowały się zbliżonym do mężczyzn poziomem w zakresie: szybkiego narastania siły globalnej i lokalnej, prędkości lokalnych ruchów, tak pojedynczych jak i wykonywanych z maksymalną częstotliwością oraz szybkości reakcji na bodźce. Podobne wyniki przedstawiciele obydwu płci uzyskali w zakresie umiejętności dokładnego odtwarzania zadanej wielkości ruchów. W bardzo zbliżony sposób u kobiet i mężczyzn układały się też wartości błędów uzyskiwane w poszczególnych zadaniach wymagających optymalnego przejawienia zdolności motorycznych, będące miarą różnych rodzajów dokładności w zakresie zdolności motorycznych (ryc. 19).
Potwierdza to dane o mniejszym dymorfizmie płciowym koordynacji ruchowej, która jest uwarunkowana wieloma specyficznymi czynnikami [Fleischman 1964; Barański 1969; Schnabel 1974; Hirtz 1977; Celikovsky i wsp. 1978; Mĕkota 1986; Osiński 1988b; Pawłowa-Starosta, Starosta 1999]. W zakresie koordynacyjnych zdolności motorycznych kobiety mogą prezentować poziom typowy dla mężczyzn, a nawet go przekraczać [Dannhauer 1977; Wyżnikiewicz-Kopp 1986; Janusz, Jarosińska 1979]. Potwierdza to także piątą hipotezę o podobnych możliwościach w zakresie przejawiania zdolności motorycznych, uwarunkowanych siłą i ruchliwością układu nerwowego u przedstawicieli obydwu płci. Natomiast nie ma podstaw do wnioskowania o istnieniu różnic w przejawianiu zdolności motorycznych u kobiet i mężczyzn w zależności od lokalnego lub globalnego charakteru zadania ruchowego.
Rycina 19 Błędy w zadaniach motorycznych wymagających maksymalnej dokładności (mężczyźni n = 176; kobiety n = 84)
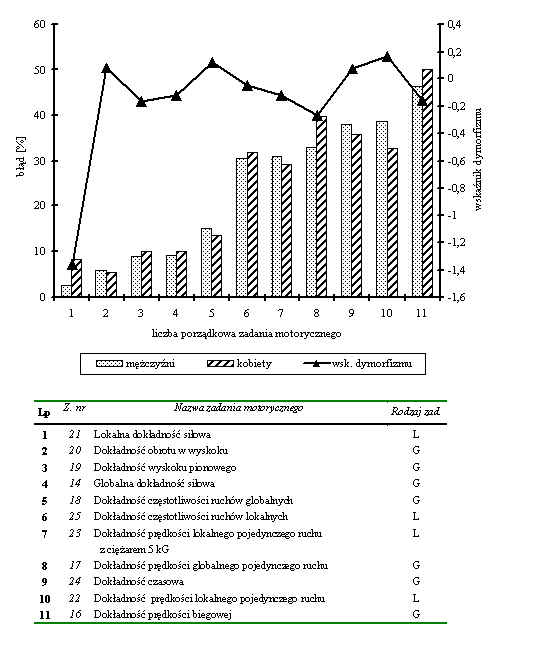
Wydaje się, że mniejsze wartości zróżnicowania płciowego stwierdzone w badaniach własnych były efektem podobnej aktywności ruchowej badanych kobiet i mężczyzn. Różnice w zakresie aktywności przedstawicieli obydwu płci stanowią jeden z zasadniczych wyznaczników dymorfizmu płciowego [Yandell, Spirduso 1981].
Dążąc do realizacji piątego celu, stwierdzono na podstawie badań własnych, że poziom zdolności motorycznych u kobiet i mężczyzn był zróżnicowany w zależności od ich umiejscowienia w strukturze tychże zdolności. Posługując się wskaźnikiem dymorfizmu, określono, że praktycznie nie było różnic w obrębie koordynacyjnych zdolności motorycznych (ryc. 20).
Określono natomiast pewne zróżnicowanie w zakresie kondycyjnych zdolności motorycznych, szczególnie związanych z przejawianiem maksymalnej siły statycznej i skoczności. Natomiast nie zaobserwowano wyraźnych różnic związanych z lokalnym lub globalnym przejawianiem się zdolności motorycznych. Ten stan sugeruje potrzebę odmiennego kształtowania kondycyjnych i koordynacyjnych zdolności motorycznych u osób obu płci, przy wykorzystaniu środków treningowych zarówno o globalnym, jak i lokalnym oddziaływaniu na ćwiczących. Jest to kolejne zagadnienie o dużym znaczeniu, które dotychczas traktowano marginalnie w teorii i praktyce wychowania fizycznego, a szczególnie treningu sportowego. Według Starosty [1995b] w działalności praktycznej dominuje tzw. „męski” model treningu, jako bardziej skuteczny i sprawdzony. Wydaje się, że w tak prowadzonym procesie rozwoju zdolności motorycznych należy uwzględnić zarówno podobne możliwości koordynacyjne kobiet i mężczyzn, jak też inną strukturę możliwości energetycznych – szczególnie typowe dla kobiet, ograniczone możliwości w zakresie beztlenowych procesów energetycznych.
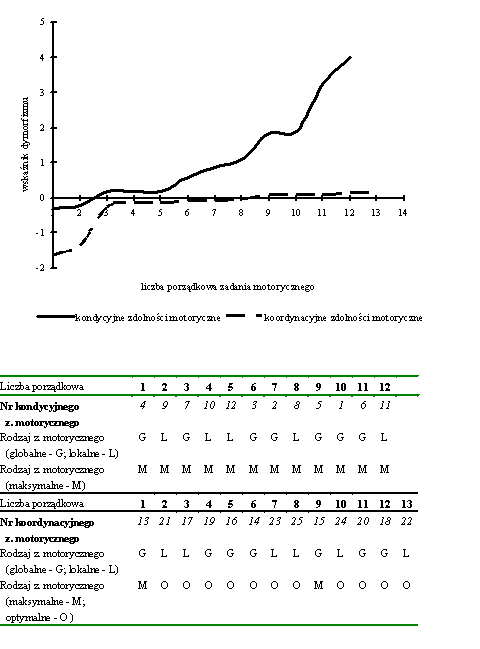
W pracy przedstawiono wyniki badań związków występujących pomiędzy globalnymi i lokalnymi przejawami kondycyjnych oraz koordynacyjnych zdolności motorycznych. Stanowi to całkowicie nowy aspekt badań nad przejawami zdolności motorycznych. Jego znaczenie jest bardzo ważne, gdyż niejednokrotnie stosuje się zestawy testów, dążąc do wszechstronnego określenia przejawów zdolności motorycznych. Często w nadziei, że przyniesie to lepsze rezultaty niż wykorzystanie ich mniejszej liczby a nawet jednego testu do kompleksowej oceny możliwości motorycznych. Nie wiadomo jednak, czy takie postępowanie nie prowadzi do niewłaściwych wniosków. Nie zawsze rozwój zdolności motorycznych na poziomie lokalnym jest podobny do ich przejawiania się w sposób globalny. Przyczyną różnic mogą być ograniczenia energetyczne lub, co jest bardziej prawdopodobne, odmienne mechanizmy informacyjne, związane ze sterowaniem ruchami na lokalnym i globalnym poziomie. Problematyka ta nie była rozważana w piśmiennictwie pomimo swojego niebagatelnego znaczenia. Równocześnie jednak istnieje szereg zaleceń metodycznych o konieczności rozwijania lokalnych i globalnych przejawów zdolności motorycznych oraz ich uwarunkowań funkcjonalnych.
Wyniki uzyskane w pracy sugerują potrzebę odmiennego traktowania globalnych i lokalnych przejawów kondycyjnych oraz koordynacyjnych zdolności motorycznych. Jest to bardzo ważne dla dalszych badań nad strukturą zdolności motorycznych, szczególnie w aspekcie doboru zadań testowych oraz praktyki wychowania fizycznego i sportu w ustalaniu zakresu stosowanych środków treningowych.
W badaniach własnych dążono też do określenia możliwości dokładnego osiągania określonych wartości rozmaitych zdolności motorycznych. Większość tych zagadnień ma bardzo duże znaczenie teoretyczne i praktyczne. Równocześnie, są to niedostatecznie szeroko zbadane obszary wiedzy o zdolnościach motorycznych. Zagadnieniami dostosowania motorycznego, które zależy od zdolności różnicowania ruchu, zajmowano się głównie od strony teoretycznej. Nieliczne wyniki badań eksperymentalnych dotyczyły pojedynczych zdolności lub ogólnych przejawów możliwości sterowania czynnościami motorycznymi. Brakuje natomiast badań nad porównaniem dokładnego przejawiania różnych zdolności motorycznych. Jest to bardzo istotne zagadnienie, ponieważ w sporcie i w życiu codziennym najczęściej niezbędne jest przejawianie ruchów w sposób optymalny, a nie maksymalny. Natomiast większość prac, w których przedmiotem badań były zdolności motoryczne, dotyczyło wysiłku o charakterze maksymalnym.
W dalszych badaniach konieczne staje się określenie możliwości przejawiania optymalnych wartości, poczynając od najmniejszego poziomu, a kończąc na zbliżonym do maksymalnego. Należy też określić wartości progowe, tzn. takie, które stanowią osobniczą granicę różnicowania intensywności przejawiania poszczególnych zdolności motorycznych. Wiedza o takich umiejętnościach jest bardzo ważna dla realnego (możliwego do wykonania) określania zasad indywidualnego dozowania intensywności wysiłku przez poszczególnych ćwiczących.
Problematyka podjęta w pracy była bardzo obszerna i złożona, dlatego też nie została w niej całkowicie zrealizowana. Wydaje się, że konieczne są dalsze obserwacje oparte na materiale badawczym, rozmaitym pod względem wieku, płci, budowy ciała i rodzaju uprawianej dyscypliny sportowej (wykonywanego zawodu) oraz zaawansowania sportowego (biegłości zawodowej). Wydaje się też, że w dalszych badaniach konieczne jest rozszerzenie zakresu stosowanych metod badawczych, zwłaszcza o pomiary energetycznych uwarunkowań zdolności motorycznych, oraz wykorzystanie wyników uzyskanych w przeprowadzonych badaniach, które w podstawowym zakresie przedstawiono w formie następujących wniosków:
Stwierdzono, że zgodnie z pierwszą hipotezą, lokalne i globalne koordynacyjne zdolności motoryczne były niezależne. Częściowe powiązania wystąpiły w obrębie lokalnych i globalnych przejawów kondycyjnych zdolności motorycznych. Chociaż stwierdzone zależności nie były zbyt silne. Wydaje się, że przyczyn należy upatrywać w czynnikach środowiskowych, związanych z istnieniem rozmaitych programów ruchów, które mogą być różne i niezależne dla lokalnych i globalnych przejawów zdolności motorycznych – szczególnie koordynacyjnych. Może to sugerować potrzebę rozwijania zdolności koordynacyjnych przy wykorzystaniu specyficznych dla nich środków treningowych, tak aby tworzyć nowe i doskonalić istniejące wzorce (programy) ruchu. Natomiast globalne i lokalne kondycyjne zdolności motoryczne może cechować pewien brak specyficzności, co powinno uprościć dobór środków wykorzystywanych do ich rozwoju, a skierowanych głównie na kształtowanie energetycznych podstaw zdolności motorycznych.
Nie znaleziono przesłanek do wnioskowania o istnieniu silnych koordynacyjnych uwarunkowań kondycyjnych zdolności motorycznych, niezależnie od globalnego lub lokalnego charakteru przejawiania się tych zdolności. Oznacza to pozytywną weryfikację drugiej hipotezy. Wydaje się, że kondycyjne zdolności motoryczne są bardziej uzależnione od czynników wrodzonych. Natomiast koordynacyjne zdolności motoryczne, niezależnie od wrodzonych możliwości układu nerwowego, są w większym stopniu uwarunkowane środowiskowo. Zatem w procesie rozwijania obydwu grup zdolności motorycznych należy wykorzystywać zadania ruchowe doskonalące procesy informacyjne, związane z przewodzeniem bodźców i tworzeniem programów ruchu oraz wymagające zaangażowania mechanizmów energetycznych.
Zgodnie z trzecią hipotezą dokładne (dozowane) odtwarzanie zadanej wielkości wybranych zdolności motorycznych zależało od ich maksymalnej wartości. Ponadto, przejawianie dokładności w rozmaitych zadaniach motorycznych było uwarunkowane poziomem zdolności kinestetycznego różnicowania ruchu i stopniem złożoności zadania ruchowego. Precyzja a zatem dokładne wykonanie ruchu zależy od możliwości zaplanowania jego wykonania i kontroli jego przebiegu oraz korygowania sposobu realizacji ruchu. Natomiast lokalny lub globalny charakter zadań ruchowych nie ma specjalnego znaczenia dla efektywności dokładnego wykonania ruchu.
Wyniki wykonania większości analizowanych zadań wymagających przejawiania kondycyjnych i koordynacyjnych zdolności motorycznych cechował znaczny stopień symetrii. Stanowiło to częściową, pozytywną weryfikację kolejnej hipotezy. Wyjątek stanowiła lokalna dokładność siłowa i dokładność lokalnej częstotliwości ruchów, które były wyraźnie asymetryczne. Prawdopodobnie, różnice w przejawianiu się zdolności motorycznych obydwu kończyn górnych wynikały z ich rozmaitej roli w wykonywaniu czynności codziennych, a zatem miały bardziej charakter wyuczony niż wrodzony. Wydaje się, że lateralizacja uzależniona od budowy układu nerwowego w mniejszym stopniu przejawia się w lokalnych zadaniach motorycznych, wymagających większej dokładności. Można przypuszczać, że ograniczenia w zakresie możliwości symetrycznego wykonywania zadań motorycznych są większe w przypadku zadań o maksymalnej niż optymalnej aktywności.
Potwierdzono także ostatnią, piątą hipotezę. Kobiety i mężczyźni uzyskali podobne wyniki w zakresie, uwzględnionych w badaniach, lokalnych i globalnych koordynacyjnych zdolności motorycznych uwarunkowanych siłą i ruchliwością układu nerwowego. Natomiast zróżnicowanie poziomu zdolności motorycznych u kobiet i mężczyzn w zależności od ich umiejscowienia w strukturze, dzielącej je na kondycyjne i koordynacyjne, sugeruje potrzebę odrębnego ich kształtowania u przedstawicieli każdej płci. Wydaje się, że pomimo istotnych ograniczeń możliwości energetycznych u kobiet nie ma różnic w stosunku do mężczyzn w zakresie tworzenia wzorców ruchu i ich wykorzystywania w złożonych koordynacyjnie zadaniach ruchowych.
* Pracę zrealizowano w ramach finansowanego przez KBN zadania: „Uwarunkowania rozwoju koordynacji ruchowej”.
Agaszin F.K. (1977) Biomechanika udarnych dviżenij. Fizkultura i Sport, Moskva.
Anioł-Strzyżewska K., Starosta W. (1984) Symmetry and Asymmetry of Movement and Their Influence on Distinctness of Kinesthetic Sensations. (In:) Genetics of Psychomotor Traits in Man, (ed.) N. Wolański, A. Siniarska. Polish Academy of Sciences, International Society of Sport Genetics and Somatology. Warszawa, s. 273–281.
Arska-Kotlińska Z., Drozdowski Z. (1995) Przejawy dymorfizmu płciowego w cechach recesywnych człowieka w populacji i grupach o ukierunkowanej aktywności ruchowej. Materiały II Krajowej Konferencji Naukowej „Problemy dymorfizmu płciowego w sporcie”, (red.) S. Socha. AWF Katowice–PTNKF, Katowice, s. 79–82.
Balsewicz W.K. (1969) K problemie fizkulturno-sportivnoj orientacii. „Teorija i Praktika Fiziczeskoj Kultury”, no. 6, s. 31–33.
Balsewicz W.K. (2000) Ontokinezjologija Czełowieka. „Teorija i Praktika Fiziczeskoj Kultury”, Moskva.
Barański A. (1969)Próba klasyfikacji nominalnych znamion motoryczności człowieka. Wychowanie Fizyczne i Sport”, 82, nr 3, s. 66–76.
Bartosiewicz G., Wit A. (1985) Skoczność czy moc? „Sport Wyczynowy”, nr 6, s. 7–14.
Bernstein N.A. (1947) O postrojenii dwiżenij. Medgiz, Moskva.
Bernstein N.A. (1966) Oczerki po fizjologii dwiżenij i fizjologii aktiwnosti. Medicina, Moskva.
Bernstein N.A. (1975) Bewegungphysiologie. Sportmedizinische Schriftenreihe, der DHfK, Band 9, Leipzig.
Bernstein N.A. (1991) O lowkosti i jej razwitiji. Fizkultura i Sport, Moskva.
Blume D.D. (1978a) Zu einigen wesentlichen theoretischen Grundpozitionen für die Untersuchung der koordinativen Fähigkeiten. „Theor. Prax. Körperkult.“, Nr. 1; s. 29–36.
Blume D.D. (1978b) Grundsatze und methodische Maßnamen zur Schulung koordinativen Fähigkeiten. „Theor. Prax. Körperkult.“, 2, s. 3–11.
Blume D. D. (1981) Kennzeichnung koordinativen Fähigkeiten und Möglichkeiten ihrer Herausbildung in Trainingprozess. „Wissenschaftliche Zeitschrift der DHfK“, 22/1, Leipzig, s. 17–41.
Blume D.D., Zimmermann K. (1987) Koordinativen Fähigkeiten. (In:) Bewegungslehre – Sportmotorik, (Hrsg.) K. Meinel, G. Schnabel. Volk und Wissen, Berlin, s. 242–247.
Bracha Cz., Chromiński Z. (1979) Trafność i rzetelność testu sprawności fizycznej. „Kultura Fizyczna”, nr 10, s. 7–9.
Celichowski J. (1995) Jednostki ruchowe (przebieg skurczu, męczliwość, rekrutacja w czasie skurczu mięśnia). Monografie nr 318, AWF w Poznaniu, s. 147–148.
Celikovsky S. i wsp. (1978) Woprosy antropomotoriki w fiziczeskom wospitaniji i sportie. Karlow Uniwersitet, Praga.
Celikovsky S. i wsp. (1979) Antropomotorika. Statni pedagogicke nakladatelstvi. Praha.
Clarke D.H. (1968) Force-time curves of voluntary muscular contraction at varying tensions. „Res. Quart.”, 39, s. 900–907.
Clarke D.H., Henry P.M. (1961) Neuromotor specifity and increased speed from strength development. „Res. Quart.”, vol. 32, no. 3, s. 315–325.
Combee F.Z. (1970) A factoral analysis of motor coordination. (In:) Contemporary readings in sport psychology, (ed.) W. P. Morgan. Springfield, s. 187–205.
Czajkowski Z. (1993) Od zwinności do zdolności zbornościowych. „Sport Wyczynowy” nr 3–4, s.18–33.
Czchaidze L.W. (1962) Koordynacja ruchów dowolnych i powstawania nawyków ruchowych człowieka w świetle ogólnych zasad sterowania i układów sterowanych. „Wychowanie Fizyczne i Sport” t. VI, z. 2.
Czchaidze L.W. (1966) Problemy centralnoj riegulacji i biomechaniczeskoj struktury dwigatielnych nawykov czelowieka. „Teorija i Praktika Fiziczeskoj Kultury”, no. 10, s. 11–13.
Czchaidze L.W. (1970) Ob uprawleniji dwiżeniami czełowieka. Fizkultura i Sport, Moskwa.
Czebyszewa W.W. (1969) Psichologija trudowogo obuczenija. Proizweszczenije. Moskwa.
Dannhauer H. (1977) Geschlecht und Persönlichkeit. Eine Intersuchung zur psychischen Geschlechts Differenzierung in der Ontogenese. Veb. Deutscher Verlag der Wissenschaften, Berlin.
Denisiuk L., Milicerowa H. (1969) Rozwój sprawności motorycznej dzieci i młodzieży w wieku szkolnym. PZWS, Warszawa.
Djaczkov W.M. (1975) O sensornoj osnowie techniko-takticzeskogo masterstwa. „Teorija i Praktika Fiziczeskoj Kultury”, no. 7, s. 21–23.
Donskoj D.D., Zaciorski W.M. (1979) Biomechanika. Fizkultura i Sport, Moskwa.
Drabik J. (1982a) Dymorfizm płciowy a wyniki sportowe. „Wychowanie Fizyczne i Sport”, 1, s. 71–73.
Drabik J. (1982b) Stronne zróżnicowanie sprawności siłowo-szybkościowej kończyn górnych i dolnych dzieci i młodzieży. „Wychowanie Fizyczne i Sport” nr 3–4.
Drabik J. (1983) Wybrane zdolności koordynacyjne u osób w różnym wieku z uwzględnieniem symetrii i asymetrii ruchu. Zeszyty Naukowe AWF w Gdańsku, nr 7, s. 255–262.
Drabik J. (1984) Sprawność fizyczna dzieci w wieku 7–15 lat w świetle symetrii i asymetrii funkcjonalnej. „Wychowanie Fizyczne i Sport”, nr 3-4, s. 57–71.
Drozdowski S. (1976) Z badań związków mocy i szybkości u studiujących wychowanie fizyczne. Monografie nr 56, AWF w Poznaniu, s. 229–231.
Drozdowski S. (1980) Rytm wybranych cech sprawności fizycznej w czasie czteroletnich studiów wychowania fizycznego. Monografie nr 126, AWF w Poznaniu, s. 17–30.
Drozdowski S., Drozdowski Z. (1974) Pomiar sprawności fizycznej ogólnej i specjalnej. Skrypty nr 24, AWF w Poznaniu, s. 9–35.
Drozdowski Z. (red.) (1980) Dymorfizm płciowy i jego przejawy w sporcie. Monografie nr 140, AWF w Poznaniu.
Drozdowski Z. (1981) Biologiczne wyznaczniki maksymalnej sprawności fizycznej. I Kongres Naukowy Kultury Fizycznej i Sportu. Materiały i dokumenty. GKKFiS, Warszawa, s. 261–265.
Drozdowski Z., (1982) O potrzebie i celowości kompleksowego ujęcia stopnia dymorfizmu płciowego w budowie ciała. Roczniki Naukowe AWF w Poznaniu, z. 31, PWN, Warszawa –Poznań.
Dumitrescu V. (1975) Coeficienti de determinare partiali si multipli. „Educatie Fizica si Sport”, nr 9, s. 1–6.
Dżamgarov T.T., Puni A.C. (1979) Psichołogija fiziczeskogo vospitanija i sporta. Fizkultura i Sport, Moskva, ss. 144.
Eurofit (1989) Europejski test sprawności fizycznej. AWF Kraków.
European Test of Physical Fitness (1988) Council of Europe for the Development of Sport. Rome.
Farfel V. (1960) Fizjologija Sporta. Fizkultura i Sport, Moskva, ss. 384.
Farfel V. (1983) Bewegungssteuerung im Sport. Berlin.
Fidelus K. (1961) Niektóre biomechaniczne zagadnienia koordynacji ruchów w świetle badań prądów czynnościowych. „Wychowanie Fizyczne i Sport”, nr 5, nr 4.
Fidelus K. (1972) Próba ustalenia podstawowych czynników motorycznych wpływających na rezultat sportowy. Roczniki Naukowe AWF, Warszawa, t. 16, PWN, Warszawa.
Fidelus K. (1983) Procesy koordynacji ruchu a jego nauczanie. Zeszyty Naukowe AWF we Wrocławiu, nr 33, s. 27–28.
Fidelus K., Skorupski L. (1970) Wielkość momentów sił mięśniowych w poszczególnych stawach u zawodników różnych dyscyplin sportu. Materiały Sympozjum Techniki Sportowej. Sport i Turystyka, Warszawa, s. 128–141.
Figura F. i wsp. (1991) Assessment of static balance in children. „The Journal of Sports Medicine and Physical Fitness”. vol. 31, no. 2, s. 235–242.
Filin W.P. (1987) Junoszeski sport. Fizkultura i Sport, Moskwa.
Filipowicz W.I., Turewski I.N. (1977) O principach sportiwnoj orientaciji detej i podrostkow w swjazi s wozrostnoj izmiencziwostju struktury dwigatelnych sposobnostiej. „Teorija i Praktika Fiziczeskoj Kultury”, no. 4, s. 39–43.
Fleishman F.A. (1964) The structure and measurement of physical fitness. Prentice-Hall, Englewood Cliffs N.J.
Fleishman F.A. (1972) Structure and measurement of psychomotor abilities. (In:) The Psychomotor Domain Movement Behavior, (ed.) R.N. Singer. Fitness, Philadelphia.
Furdal S. (1989) Modelowanie statystyczne w badaniach empirycznych. Prace i Materiały. Tom XXX. Instytut Sportu, Warszawa, ss. 388.
Geblewiczowa M. (1965) Szybkość reagowania, szybkość ruchów człowieka. INKF, Warszawa.
Geblewiczowa M. (1968) Badania eksperymentalne nad siłą, wytrzymałością i szybkością. „Wychowanie Fizyczne i Sport”, nr 1, s. 15–26.
Geblewiczowa M. (1973) Badania nad szybkością ruchów człowieka. Studia i Monografie AWF Warszawa, nr 6.
Geblewiczowa M., Ogórek J. (1960) Czas reakcji prostych i różnicowych w zależności od płci. „Wychowanie Fizyczne i Sport”, nr 1, s. 103–108.
Gellersztejn S.G. (1958) Czuwstwa wremieni i skorost dwigatielnoj reakcji. Medicina, Moskwa.
Gilewicz Z. (1964) Teoria wychowania fizycznego. Sport i Turystyka, Warszawa.
Golema M. (1981) Biomechaniczne badania regulacji równowagi u człowieka. Studia i Monografie AWF we Wrocławiu, nr 2.
Golema M. (1987) Stabilność pozycji stojącej. Studia i Monografie AWF we Wrocławiu, zeszyt nr 17.
Golema M., Jaśkiewicz G. (1986) Próba wykorzystania modelu matematycznego jako metody dla oceny właściwości motorycznych sportowców w procesie utrzymania równowagi. „Wychowanie Fizyczne i Sport”, nr 1, Warszawa.
Górgól A. (1993) Körperhaltung bei sechs-jährigen Kindern beeinflusst.3rd International Conference „Sport Kinetics’93“, Poznań – Warszawa.
Greń J. (1976) Statystyka matematyczna. Modele i zadania. PWN, Warszawa.
Grottel K., Celichowski J. (1992) Organizacja mięśnia i sterowanie ruchem. Część I: Budowa i czynność jednostek ruchowych. Podręczniki nr 43, AWF w Poznaniu.
Grottel K., Krutki P. (1996) Organizacja mięśnia i sterowanie ruchem. Część II: Sterowanie ruchem. Podręczniki nr 46, AWF w Poznaniu; Wyd. Nauk. PWN, Poznań – Warszawa.
Guilford J.P. (1958) A system of the psychomotor abilities. „American Journal Psychology”, 71, s. 164–174.
Gundlach H. (1968) Systembeziehungen körperlicher Fähigkeiten und Fertigkeiten. „Theor. Prax. Körperkult.“, Nr. 7.
Gundlach H. (1970) O systemie zależności pomiędzy zdolnościami i umiejętnościami fizycznymi. (W:) Sympozjum teorii techniki sportowej. Sport i Turystyka, Warszawa, s. 185–194.
Harre D. (1985) Grundlage und Methodik der Ausbildung koordinativer Fähigkeiten. „Theor. Prax. Körperkul.“, Nr. 2, s. 198–205.
Havli ek I. (1972) Overanie platnosti testov pohybovej vykonnosti metodou faktorovej analizy. „Teorie a Praxe Tělesnoj Vỳchovy“, 20, s. 70–78.
Henry P.M. (1960) Factorial structure of speed and static strength in a lateral arm movement. „Res. Quart.”, vol. 31, no. 3, s. 440–447.
Henry P.M., Whitley J.D. (1960) Relationships between individual differences in strength, speed, and mass in arm movement. „Res. Quart.”, vol. 31, no. 1 s. 24–33.
Hirtz P. (1976) Untersuchungen zur Entwicklung koordinativen Leistungsvoraussetzungen bei Schulkindern. „Theor. Prax. Körperkult.“, Nr. 4, s. 283–289.
Hirtz P. (1977) Struktur und Entwicklung koordinativen Leistungsvoraussetzungen bei Schulkindern. „Theor. Prax. Körperkult.“, Nr. 7 s. 503–510.
Hirtz P. (1985) Koordinativen Fähigkeiten im Schulsport. Volk und Wissen, Berlin, s. 152.
Hirtz P. (1986) Entwicklung koordinativen Fähigkeiten im Schulalter und moglichkeiten ihrer Vervollkommung. (W:) Motoryczność dzieci i młodzieży – aspekty teoretyczne oraz implikacje metodyczne, (red.) J. Raczek. Cz. 1. AWF Katowice, s. 75–83.
Hirtz P. (1989) Phänomen koordinativen Fähigkeiten. „Theor. Prax. Körperkult.“, Beiheft 2, Berlin, s. 30–33.
Hirtz P., Starosta W. (1990) Entwicklungstrends in der Erforschung der Bewegungskoordination im Sport. (In:) Bewegungskoordination im Sport, (Hrsg.) W. Starosta. Internationale Gesselschaft für Sportmotorik, Warszawa – Gorzów Wlkp., s. 11–27.
Hirtz P., Starosta W. (1991) Kierunki badań koordynacji ruchowej w sporcie. „Antropomotoryka”, nr 5, s. 69–82.
Hirtz P., Starosta W. (1994) Kinastesie und Sport. 3rd International Conference „Sport Kinetics’93“, Poznań – Warszawa.
Hirtz P., Thomas S. (1977) Zur Entwicklung und Struktur koordinativ – motorischer Leistungsvoraussetzungen von Teilnehmern am außerordentlichen Sport. „Theor. Prax. Körperkult.", Nr. 3.
Hirtz P., Kirchner G., Pöhlmann R. (1994) Sportmotorik (Grundlagen, Anwendungen und Grenzgebiete). Psychomotorik in Forschung und Praxis, Band 22, Universität Gesamthochschule, Kassel.
Hotz A., Weineck J. (1983) Optimales Bewegungslernen. Erlangen.
Huxley A.F., Simmons R.M. (1972) Mechanical transients and the origin of muscular force. „Symp. Quant. Biol.”, 37, s. 669–680.
Ilin E.P. (1976) Struktura psichomotornych sposobnostiej. „Psichomotorika”, 2, s. 4–22.
Instrukcja obsługi systemu badań posturograficznych (1993). Zakład Teorii Sportu IWF w Gorzowie Wlkp.
Janusz A., Jarosińska A. (1979) Współzależność pomiędzy cechami morfologicznymi i motorycznymi dzieci w wieku 9–12 lat. „Mat. i Prace Antrop.”, nr 6.
Janusz A., Jarosińska A. (1981) Związki między sprawnością fizyczną a budową ciała dzieci wrocławskich w wieku pokwitania. „Mat. i Prace Antrop.”, nr 96, s. 39–53.
Jarosz E., Wolański N. (1967) Podobieństwo czucia proprioreceptywnego u dzieci i ich rodziców (wyniki badań rodzinnych). „Wychowanie Fizyczne i Sport”, nr 11.
Jaskólska A. (1998) Przebieg zmian narastania siły i relaksacji u ludzi po wysiłkach o różnej charakterystyce. Monografie nr 326, AWF w Poznaniu.
Jaskólska A., Jaskólski A., Adach Z., Nieścieruk-Szafrańska B. (1987) Wpływ uprawiania sportu na wielkość różnic w wydolności beztlenowej kobiet i mężczyzn. „Wychowanie Fizyczne i Sport”, nr 4, s. 41–47.
Kasa J. (1983) Struktura pohybowych schopnosti a zrucnosti. „Trener”, nr 9, s. 1–16.
Kasa J., Belej M. (1994) Die Entwicklung der Anthropomotorik als Eines Wissenschaftlichen Faches in der Slowakei. 3rd International Conference „Sport Kinetics’93”, Poznań – Warszawa.
Komor A. (1983) Cybernetyczne modele identyfikacji parametrów koordynacji ruchów. Zeszyty Naukowe AWF we Wrocławiu, nr 33.
Korenberg W.B. (1979) Osnowy kaczestviennogo biomechaniczeskogo analiza. Fizkultura i Sport, Moskwa, s. 55–58.
Kossov B.B. (red.) (1973) Poznawatelnyje processy u sportsmenow. T. I. Sensornaja osnowa sportiwnogo masterstwa. Wsiesojuznyj Nauczno–Issledovatielnyj Institut Fizi eskoj Kultury, Moskva.
Kossov B.B. (red.) (1976) Poznawatelnyje processy u sportsmenow. T. II. Samokontrol i masterstwo w sporte. Wsiesojuznyj Nauczno–Issledowatielnyj Institut Fizi eskoj Kultury, Moskva.
Kruczalak E. (1979) Czynniki warunkujące poziom wyników sportowych w biegach na 100 i 200 m oraz model szkolenia biegaczy na krótkie dystanse. Wydawnictwo Monograficzne AWF w Krakowie, nr 14.
Krüger H. (1989) Zur Problematik der Wechselbeziehungen zwissen Information und Energie bei der Realisierung motorischer Handlungen. „Theor. Prax. Körperkult.", Beiheft 2, s. 110–112.
Krüger H., Zimmermann K. (1983) Koordinative Fähigkeitentwicklung und Technikschulung bei jungen Sportlern. „Theor. Prax. Körperkult.“, Nr. 11.
Kuhn W. (1987) Zum Problem des kontralateralen Transfers. Köln.
Kulig K., Suska M., Zawadzki J. (1978) Badania nad koordynacją szybkich ruchów przedramienia. „Wychowanie Fizyczne i Sport”, nr 2, Warszawa.
Kuzniecov W.W. (1975) Specjalnaja siłovaja podgotovka sportsmena. Izd. „Sovietskaja Rossija”, Moskwa, s. 122–133.
Leonov O.I. (1973) Czuwstwo tempa u konkobeżcew raznoj kwalifikaciji. (W:) Poznawatelnyje processy u sportsmenow. T.I. Moskwa.
Lienert G.A. (1969) Testaufbau und Testanalyse. Verlag J. Beltz, Berlin.
Ljach W.I. (1976) Issledowanije łowkosti detej i podrostkow w podwiżnych igrach (Autoreferat dissertaciji kandidata pedagogiczeskich nauk). Moskwa.
Ljach W.I. (1983) Ponjatja koordinacjonnyje sposobnosti i łowkost. „Teorija i Praktika Fiziczeskoj Kultury”, no. 8.
Ljach W.I. (1987) O klasifikaciji koordinacjonnych sposobnostiej. „Teorija i Praktika Fiziczeskoj Kultury”, no. 7, s. 28–30.
Ljach W.I. (1988) Ważniejszyje dla raznych widow sporta koordynacjonnyje sposobnosti i ich znaczenije w techniczeskom i technikatakticzeskom sowierszowanii. „Teorija i Praktika Fiziczeskoj Kultury”, no. 2.
Ljach W.I. (1989) Koordynacjonnyje sposobnosti szkolnikow. Polymia, Minsk.
Ljach W.I. (1990) Senzitiwnyje periody razwitja koordinacjonnych sposobnostiej dietiej w szkolnom wozrastje. „Teorija i Praktika Fiziczeskoj Kultury”, no. 3, s. 14–24.
Ljach W.I. (1991a) Idei N.A. Bernsteina i ich razwitie w naukie i praktikie. „Teorija i Praktika Fiziczeskoj Kultury”, no. 3, s. 2–9.
Ljach W.I. (1991b) Kritieria opriedielienia koordinacionnych sposobnostjej. „Teorija i Praktika Fiziczeskoj Kultury”, no. 11.
Ljach W.I. (1993) Dinamika wzaimoswiaziej mieżdu koordinacjonnymi sposobnostiami, morfologiczeskimi priznakami, kondycjonnymi sposobnostiami i pokazatieljami razwitija psichofizjołogiczeskich funkcjej u dietej. (W:) Izbrannyje aspekty sportiwnoj motoriki, (red.) W. Starosta, N. Pristupa. Brest, s. 83–91.
Ljach W.I. (1995) Przygotowanie koordynacyjne w zespołowych grach sportowych. An International Conference on Science in Sports Team Games, (ed.) J. Bergier. Instytut Wychowania Fizycznego i Sportu, Biała Podlaska, s. 153–167.
Ljach W.I. (2001) The structure of coordinational Motor Abilities (CMA) in athletes and schoolchildren and its dependence on various factors. (In:) Motor Coordination in Sport and Exercise. Atti Convegno di Studi, Bologna, s. 107–118.
Ljach W., Starosta W. (2001) The influence of genetic and environmental conditions on the variability of the level of selected co-ordination abilities of children. Studies in Physical Culture and Tourism, Vol. VIII, University School of Physical Education in Poznań, s. 127–138.
Ljach W., Mynarski W., Raczek J. (1995) Biopsychiczne predyspozycje koordynacyjnych zdolności motorycznych – przegląd badań w piśmiennictwie rosyjskojęzycznym. „Antropomotoryka”, nr 12-13, s. 83–106.
Ljach W., Witkowski Z., Żmuda W. (2000) Struktura koordynacyjnych zdolności motorycznych i kontrola koordynacyjnego przygotowania wyselekcjonowanych piłkarzy nożnych na etapie szkolenia specjalistycznego. (W:) Efektywność systemów szkolenia w różnych dyscyplinach sportu. Materiały konferencji naukowo-metodycznej w Spale. Warszawa, s. 88–93.
Łomov B.F., Kossov B.B., Konopkin O.A. (1976) Teoreticzeskije problemy samokontrola i uprawlenija sportiwnoj dejatelnostju. (W:) Poznawatelnyje processy u sportsmenow, (red.) B.B. Kossov. Wsiesojuznyj Nauczno-Issledowatielnyj Institut Fizi eskoj Kultury, Moskwa.
Mattausch W.D. (1973) Zu einigen Problemen den begrifflichen fizirung den konditionellen und koordinativen Fähigkeiten. „Theor. Prax. Körperkult.“, Nr. 9.
Matwiejew Ł.P. (1977) Osnowy sportiwnoj trenirowki. Fizkultura i Sport, Moskwa.
Mc Cloy C.M. (1960) Wpływ ćwiczeń siłowych z narastającym oporem na rozwój siły oraz jej powiązanie z poprawą zręczności ruchowej w sportach i lekkiej atletyce. „Wychowanie Fizyczne i Sport”, t. IV, 1.
Meinel K. (1960) Bewegungslehre. Volk und Wissen Volksseigener Verlag, Berlin.
Meinel K. (1967) Motoryczność ludzka. Sport i Turystyka, Warszawa, ss. 466.
Meinel K., Schnabel G. (Hrsg.) (1998) Bewegungslehre – Sportmotorik. Sportverlag, Berlin.
Mero A., Luhtanen P., Viitasalo J.T., Komi P.V. (1981) Relationship between the maximal running velocity, muscle fiber characteristics, force production and force relaxation of sprinters. „Scand. J. Sport Sci.”, 3, s. 16–22.
Mĕkota K. (1972) Struktura lidskie motoriky – metody, nektere vysledky a perspektivy vyskumu. Acta Universitatis Palackianae Olomoucensis, 40, Praha, s. 25–54.
Mĕkota K. (1982) K pojeti, strukture a testovani koordina nich schopnosti – shrnuti literarnich poznatki. Gymnica XII. Acta Universitatis Palackianae Olomoucensis, Praha, s. 5–18.
Mĕkota K. (1986) Zur diagnostik koordinativen Fähigkeiten bei Kinder und Jugendlichen. (W:) Motoryczność dzieci i młodzieży – aspekty teoretyczne oraz implikacje metodyczne, (red.) J. Raczek. AWF Katowice, s. 61–73.
Mĕkota K., Blahus P. (1983) Motoricke testy telesne vychovy. SPN, Praha. Mĕkota K., Kovař R., Stepnicka J. (1988) Antropomotorika II. Univerzita Palackeho v Olomounci, Praha.
Mieńszikov B.Ja. (1966) O sposobnosti differencirovat vremia, prostranstvo i stepień myszecznogo naprażenija. „Teorija i Praktika Fiziczeskoj Kultury”, no. 4.
Milner-Brown H.S., Stein R.B. (1973) Changes in firing rate of motor units during linearly changing voluntary contractions. „J. Physiol.”, 230, s. 371–390.
Mleczko E. (1991) Przebieg i uwarunkowania rozwoju funkcjonalnego dzieci krakowskich między 7 a 14 rokiem życia. Wydawnictwo Monograficzne AWF w Krakowie, nr 44.
Mleczko E. (1992) Przegląd poglądów na temat motoryczności człowieka. „Antropomotoryka”, nr 10, s. 109–140.
Mleczko E. (1993) Zróżnicowanie środowiskowe a poziom i dynamika rozwoju funkcjonalnego dzieci krakowskich między 7 a 14 rokiem życia. „Antropomotoryka”, nr 10, s. 57–104.
Morecki A., Ekiel J., Fidelus K. (1971) Bionika ruchu. PWN, Warszawa.
Mynarski W. (1986) Właściwości motoryczności ludzkiej w świetle współczesnych psychologicznych teorii zdolności. (W:) Motoryczność dzieci i młodzieży – aspekty teoretyczne oraz implikacje metodyczne, (red.) J. Raczek. AWF Katowice, s. 86–89.
Mynarski W. (2000) Struktura wewnętrzna zdolności motorycznych dzieci i młodzieży w wieku 8–18 lat. AWF Katowice.
Naglak Z. (1979) Trening sportowy. Teoria i praktyka. PWN, Warszawa–Wrocław.
Neumeier A. (1983) Sportmotorische Tests in Unterricht und Training. Verlag K. Hoffmann, Schorndorf.
Nowacki Z., Siemieński A. (1990) Siła kończyn górnych a dokładność manipulacji. Materiały Ogólnopolskiej Konferencji Biomechaniki. AWF Gdańsk, s. 351–360.
Osiński W. (1977) Analiza związków między szybkością lokomocyjną a siłą, mocą i skocznością u chłopców. Roczniki Naukowe AWF w Poznaniu, z. 26, PWN, Warszawa–Poznań, s. 91–108.
Osiński W. (1981) Wysokość i masa ciała a czas reakcji i dyspozycje motoryczne u kobiet i mężczyzn. Roczniki Naukowe AWF w Poznaniu, z. 30, PWN, Warszawa–Poznań, s. 97–105.
Osiński W. (1985) Badania nad wartością pomiaru wybranych zdolności motorycznych. „Przegląd Antropologiczny”, nr 1-2, s. 39–55.
Osiński W. (1988a) Wielokierunkowe związki zdolności motorycznych i parametrów morfologicznych. Badania dzieci i młodzieży wielkomiejskiej z uwzględnieniem stratyfikacji społecznej. Monografie nr 261, AWF w Poznaniu, s. 38–73.
Osiński W. (1988b) Zależność między zdolnościami motorycznymi a cechami, wskaźnikami i komponentami ciała u dzieci i młodzieży. „Wychowanie Fizyczne i Sport”, nr 4, s. 11–28.
Osiński W. (1990) Uwagi na tle definicji i klasyfikacji podstawowych pojęć charakteryzujących motoryczność człowieka. „Antropomotoryka”, nr 3, s. 3–8.
Osiński W. (red.) (1993) Motoryczność człowieka – jej struktura, zmienność i uwarunkowania. Monografie nr 310, AWF w Poznaniu.
Osiński W. (2000) Antropomotoryka. Wyd. I. Podręczniki nr 49, AWF w Poznaniu.
Osiński W., Swornowski P. (1983) Analiza przejawów dymorfizmu płciowego w obrębie właściwości motorycznych. „Przegląd Antropologiczny”, t. 49, z. 1–2, s. 27–39.
Osiński W., Jarecki G., Poprawski B., Strzelczyk R., Wachowski E., Winkler A. (1978) Z badań związków szybkości biegowej. Monografie nr 115, AWF w Poznaniu, s. 499–508.
Osiński W., Szeklicki R., Kowalczyk J., Biernacki J., Kusy J., Maciaszek J., Rauk M. (1994) Sex Differentiation in Coordinative Motor Abilities. 3rd International Conference „Sport Kinetics’93”, Poznań–Warszawa.
Ozierov V.P. (1983) Psichomotornyje razvitije sportsmenov. Sztiinca, Kisziniev.
Ozolin I.G. (1970) Sowremiennaja sistema sportiwnoj trenirowki. Fizkultura i Sport, Moskwa.
Pańczyk J. (1981) Rozmiary dymorfizmu płciowego w rozwoju cech motorycznych u uczniów w wieku 9–18 lat. „Kultura Fizyczna”, nr 1, s. 10–11.
Pawłowa-Starosta T., Starosta W. (1999) Sex dimorphism within the selected somatic traits and motor abilities of girls and boys aged 10–11. Proceedings I of 6 Sport Kinetics Conference ’99 „Theories of Human Motor Performance and their Reflections in Practice”, Ljubljana 1–4.09.1999, (ed.) W. Strojnik, A. Ušaj. Faculty of Sport University of Ljubljana, Ljubljana, s. 265–268.
Pilicz S. (1986) Analiza czynnikowa struktury sprawności fizycznej. (W:) Motoryczność dzieci i młodzieży – aspekty teoretyczne oraz implikacje metodyczne, (red.) J. Raczek. AWF Katowice, s. 171–176.
Pilicz S., Witczak T. (1977) Struktura szybkości w zależności od płci, wieku i zaawansowania ruchowego badanych. „Wychowanie Fizyczne i Sport”, nr 1, s. 63–73.
Popinigis J. (1997) O powinnościach biochemii w stosunku do lekkiej atletyki. (W:) Lekkoatletyka – biegi. AWF Gdańsk.
Popow J., Stepanenko O. (1978) Sprint: uprażnienija so sztangoj. „Legkaja Atletika”, no. 3, s. 20–21.
Pöhlmann R. (1986) Motorisches Lernen. Berlin, ss. 270.
Pöhlmann R., Kirchner G., Wohlgefahrt K. (1979) Der Psychomotorische Fähigkeitskomplex seine Kennziechen und seine Vervollkommung. „Theor. Prax. Körperkult.”, 11, s. 898–907.
Prus G. (2000) Wytrenowalność zdolności motorycznych kobiet. AWF Katowice.
Prus G., Szopa J. (1997) Adaptabilność wybranych zdolności motorycznych u chłopców między 12 a 15 rokiem życia: rezultaty eksperymentu „trening – detrening – retrening”. „Antropomotoryka”, nr 16, s. 27–43.
Przewęda R. (1973) Rozwój somatyczny i motoryczny. WSiP, Warszawa.
Puni A.C. (1955) Kinesteticzeskoje prostrastwiennoje rozliczenije w sportiwnoj diejatielnosti. (W:) Probljemy wosprijatja prostranstwa i prostranstwiennych otnoszenij, (red.) B. Ananiew, B. Łomow. Izd. APN RSFSR, Moskva, s. 139–146.
Puni A.C. (1957) O czuvstviennych i logiczeskich komponentach dvigatielnoj dejatielnosti sportsmenov. Materiały sovieszczanija po psichołogii. Izd. APN RSFSR, Moskwa.
Puni A.C. (1963) Kinesteticzeskoje prostrastwiennoje rozliczenije w sportiwnoj diejatielnosti. Leningrad.
Puni A.C. (1967) Pamięć ruchowa i działalność ruchowa. „Kultura Fizyczna”, nr 11, s. 526–529.
Puni A.C. (1984) Psichołogija. Fizkultura i Sport, Moskva.
Raczek J. (1986) Motoryczność człowieka, poglądy, kontrowersje i koncepcje. (W:) Motoryczność dzieci i młodzieży – aspekty teoretyczne oraz implikacje metodyczne, (red.) J. Raczek. Cz. I. AWF Katowice.
Raczek J. (1987) Motoryczność człowieka w świetle współczesnych poglądów i badań. „Wychowanie Fizyczne i Sport”, nr 1, s. 5–25.
Raczek J. (1990) Koordinativ–motorische Vervollkommung und sportmotorische Lernerfolge im Sport Unterricht und Nachwuchstraining. „Leistungssport”, Nr. 5.
Raczek J. (1991a) Koordynacyjne zdolności motoryczne: Podstawy teoretyczne, empiryczne i znaczenie w sporcie. „Sport Wyczynowy”, nr 5/6.
Raczek J. (1991b) Podstawy szkolenia sportowego dzieci i młodzieży. Warszawa.
Raczek J. (1993) Koncepcja strukturalizacji i klasyfikacji motoryczności człowieka. (W:) Motoryczność człowieka – jej struktura, zmienność i uwarunkowania, (red.) W. Osiński. Monografie nr 310, AWF w Poznaniu.
Raczek J. (1994) Theorie der Motorik – Entwicklungstendenzen, Grundfragen und Stellenwert im System Sportwissenschaften. 3rd International Conference „Sport Kinetics‘93”, Poznań – Warszawa.
Raczek J., Mynarski W. (1991) Z badań nad strukturą koordynacyjnych zdolności motorycznych. „Antropomotoryka”, nr 5, PWN, Kraków, s. 3–19.
Raczek J., Mynarski W. (1992) Koordynacyjne zdolności motoryczne dzieci i młodzieży. Struktura wewnętrzna i zmienność osobnicza. (W:) Studia nad motorycznością ludzką. AWF Katowice.
Raczek J., Mynarski W., Ljach W. (1998) Teoretyczno-empiryczne podstawy kształtowania i diagnozowania koordynacyjnych zdolności motorycznych. (W:) Studia nad motorycznością ludzką. AWF Katowice, s. 36–98.
Raczek J., Mynarski W., Ljach W. (2002) Kształtowanie i diagnozowanie koordynacyjnych zdolności motorycznych. Podręcznik dla nauczycieli, trenerów i studentów. AWF, Katowice.
Radzińska M., Starosta W. (2002) Znaczenie, rodzaje i struktura skoczności oraz jej uwarunkowania. Międzynarodowe Stowarzyszenie Motoryki Sportowej, Instytut Kultury Fizycznej, Gorzów Wlkp.
Regner K. (1973) Silova obratnost. „Teorie a Praxe Tělesnoj Vỳchovy”, 11, s. 687–692.
Roth K. (1982) Strukturanalyse koordinativen Fähigkeiten. Limpert Verlag, Bad Homburg, s. 37–54.
Rożynek-Łukanowska W. (1979) Specyfika wydolności fizycznej kobiet. „Kultura Fizyczna”, nr 8, s. 4–8.
Rudik P.A. (1958) Psichołogija sporta. Fizkultura i Sport, Moskva.
Rutkowska-Kucharska A. (1982) Struktura koordynacji ruchów a przesłanki do uprawiania gimnastyki artystycznej. Autoreferat rozprawy doktorskiej. AWF Wrocław.
Rutkowska-Kucharska A., Bober T. (1986) Badania koordynacji ruchów. (W:) Potencjał ruchowy człowieka, (red.) T. Bober. Warszawa, s. 100–143.
Rutkowska-Kucharska A., Bober T., Golema M. (1990) Weryfikacja metod oceny manipulacją i równowagą człowieka. Materiały Ogólnopolskiej Konferencji Biomechaniki. AWF Gdańsk.
Rynkiewicz T. (1989) Związki mocy użytecznej w wybranych ćwiczeniach siłowo-szybkościowych z możliwościami specjalistycznymi kajakarzy. „Wychowanie Fizyczne i Sport”.
Rynkiewicz T. (1990a) Umiejętność regulacji mocy w ćwiczeniach z ciężarkami na przykładzie kajakarzy. (W:) W. Starosta (1990) Koordynacja ruchowa w sporcie. Warszawa – Gorzów Wlkp.
Rynkiewicz T. (1990b) Z badań nad efektami regulacji częstotliwości w ćwiczeniu z ciężarami. Materiały Ogólnopolskiej Konferencji Biomechaniki. AWF Gdańsk.
Rynkiewicz T., Starosta W. (1996) Selected aspects of strength manifestations and „strength accuracy” in advanced canoeists. (In:) The Proceedings of 4th International Scientific Conference „Sport Kinetics ’95”. Prague, s. 400–403.
Rynkiewicz T., Starosta W. (1999) Strength and strength’s precision in exercises of local and global character. Proceedings of 6th Sport Kinetics Conference ’99 „Theories of Human Motor Performance and their Reflections in Practice”, (ed.) V. Strojnik, A. Ušaj. Faculty of Sport University of Ljubljana, Ljubljana 1-4.09.1999, Ljubljana, s. 134–137.
Rynkiewicz T., Starosta W. (2000) Strength differentiation in girls and boys. „Biology of Sport”, Vol. 17, 3, s. 208–216.
Sankowski T. (1990) „Cechy” czy zdolności – rzecz o motoryczności. „Sport Wyczynowy”, nr 7–8, s. 35–41.
Schmidt R.A. (1976) Control processes in motor skills. „Exercise and Sport Sciences Review”, 4, s. 229–261.
Schmidt R.A. (1988) Motor control and learning. A behavioral emphasis. Human Kinetics, Champaign.
Schnabel G. (1968) Zur Bewegungskoordination. „Wissenschaftliche Zeitschrift der DHfK“, Nr. 1, Leipzig.
Schnabel G. (1974) Koordinativen Fähigkeiten im Sport – ihre Erfassung und zielgerichtete Ausbildung. „Theor. Prax. Körperkult.”, Nr. 7, s. 627–632.
Schnabel G., Harre D., Borde A. (Hrsg.) (1994) Trainingwissenschaft, Leistung, Training, Wettkampf. Sportverlag, Berlin.
Sekita B. (1970) Z badań nad zróżnicowaniem motorycznym prawej i lewej strony ciała u dziewcząt i chłopców w wieku 18 lat. „Kultura Fizyczna”, nr 7, s. 304–308.
Sieczenov I.M. (1986) Odruchy mózgowe. PWN, Warszawa, ss. 173.
Skład M. (1973) Rozwój fizyczny i motoryczność bliźniąt. „Mat. i Prace Antrop.”, t. 85, s. 117–138.
Socha Z. (red.) (1995) Dymorfizm płciowy w sporcie. Materiały II Krajowej Konferencji Naukowej „Problemy dymorfizmu płciowego w sporcie”, (red.) S. Socha. AWF Katowice – PTNKF, Katowice.
Sozański H., Witczak T. (1981) Trening szybkości. Sport i Turystyka, Warszawa.
Starosta W. (1975) Symetria i asymetria ruchów w sporcie. Sport i Turystyka, Warszawa, ss. 203.
Starosta W. (1978a) Nowy sposób pomiaru i oceny koordynacji ruchowej. Monografie nr 96, AWF w Poznaniu, s. 365–369.
Starosta W. (1978b) Nowy sposób pomiaru tzw. skoczności. Monografie nr 96, AWF w Poznaniu, s. 351–355.
Starosta W. (1983) Dokładność ruchu – jeden ze wskaźników przygotowania technicznego. Zeszyty Naukowe AWF we Wrocławiu, nr 33, s. 63–79.
Starosta W. (1984) Movement Co-ordination as Sport Selection Element. (In:) Genetics of Psychomotor Traits in Man, (ed.) N. Wolański, A. Siniarska. Polish Academy of Sciences, International Society of Sport Genetics and Somatology, Warszawa.
Starosta W. (1987) Znaczenie badań koordynacji ruchowej dla doskonalenia sportowego zaawansowanych zawodników. „Kultura Fizyczna”, nr 3–4.
Starosta W. (1990) Symetria i asymetria ruchów w treningu sportowym. Wyd. Instytutu Sportu, z. 15, Warszawa, s. 33–125.
Starosta W. (1993a) Koordynacja ruchowa człowieka. (W:) Motoryczność człowieka – jej struktura, zmienność i uwarunkowania, (red.) W. Osiński. Monografie nr 310, AWF w Poznaniu, s. 81–120.
Starosta W. (1993b) Symetria i asymetria ruchów człowieka. (W:) Motoryczność człowieka – jej struktura, zmienność i uwarunkowania, (red.) W. Osiński. Monografie nr 310, AWF w Poznaniu, s. 81–120.
Starosta W. (1995a) Zdolności koordynacyjne i kondycyjne w zespołowych grach sportowych. An International Conference on Science in Sports Team Games, (ed.) J. Bergier. Instytut Wychowania Fizycznego i Sportu, Biała Podlaska, s. 105–138.
Starosta W. (1995b) Sport dla kobiet czy kobieta dla sportu. Materiały II Krajowej Konferencji Naukowej „Problemy dymorfizmu płciowego w sporcie”, (red.) S. Socha. AWF Katowice – PTNKF, Katowice, s. 9–17.
Starosta W. (1998) Correlation between co-ordination and physical abilities in the theory and practice of sport training. Schriften der Deutschen Vereinigung für Sportwissenschaft, Band 98, Czwalina Verlag, Hamburg, vol. 1, s. 57–69.
Starosta W. (2001a) The importance of movement co-ordination, its structure and the hierarchy of integrant elements and physical education. (In:) Motor Coordination in Sport and Exercise. Atti Convegno di Studi, Bologna, s. 13–88.
Starosta W. (2001b) Uwarunkowania stronnego zróżnicowania dokładności ruchów u wysoko zaawansowanych zawodników uprawiających rozmaite dyscypliny sportowe. „Medycyna Sportowa”, 4, s. 161–164.
Starosta W., Bergier J. (1994) Das Problem der Symmetriezierung der Schusse aufs Tor in den Wettkämpfen vom Höchsten Rang in Fußball. Proceedings of the 3rd International Conference „Sport Kinetics '93”, (ed.) W. Osiński, W. Starosta. Academy of Physical Education in Poznań – Institute of Sport in Warszawa, s. 699–706.
Starosta W., Hirtz P. (1989) Sensitive and critical periods in development of coordination abilities in children and youths. „Biology of Sport”, Vol. 6, Suppl. 3, Warszawa, s. 276–282.
Starosta W., Radzińska M. (1997) Interdependence of the jumping ability results obtained via different measurement methods. International Conference „Sport Kinetics ’97”, Magdeburg.
Starosta W., Kos H., Sadowski J. (1992) Die Fähigkeit der Muskelkraftdifferenzierung unter Einfluss von Belastung. Gymnica XXII, Acta Universitatis Palackianae Olomoucensis. Olomouc, s. 333–339.
Starosta W., Rynkiewicz T., Kos H. (1994) Aus den Untersuchungen der Bedingungen für die Kraftdifferenzierungsfähigkeit („Kraftgenaufähigkeit“). (In:) Steuer - und Regelvorgänge der menschlichen Motorik, (Hrsg.) P. Blaser, K. Witte, Ch. Stucke. Academia Verlag, Sankt Augustin, s. 238–244.
Stokłosa H. (1998) Kształtowanie się asymetrii funkcjonalnej i morfologicznej 7–15 letnich dziewcząt i chłopców. AWF Katowice.
Strzelczyk R. (1995) Uwarunkowania rozwoju ruchowego dzieci wiejskich. Próba hierarchicznego ujęcia czynników determinujących. Monografie nr 324, AWF Poznań.
Szczepanik M. (1993) Wpływ treningu koordynacyjnego na szybkość uczenia się techniki ruchu u młodych siatkarzy. „Sport Wyczynowy”, nr 3–4, s. 41–51.
Szczęsna D., Guzman G., Miller T., Zhao J., Farokhi K., Ellemberger H., Potter J.D. (1996) The role of the four Ca2+ binding sites of troponin C in the regulation of skeletal muscle contraction.” J. Biol. Chem.”, 271, s. 8381–8386.
Szczotka F. (1969) Elementarne metody statystyki i ich zastosowania w naukach o wychowaniu fizycznym. AWF Warszawa.
Szeklicki R., Osiński W., Biernacki J., Rauk M., Kowalczyk J., Kusy K., Maciaszek J. (1995) Zróżnicowanie płciowe oraz morfologiczne i motoryczne uwarunkowania równowagi ciała. Monografie nr 318, AWF w Poznaniu, s. 7–13.
Szopa J. (1985) Zmienność ontogenetyczna, zróżnicowanie środowiskowe oraz genetyczne uwarunkowania rozwoju komponentów ciała populacji wielkomiejskiej w wieku 7–63 lata. Wydawnictwo Monograficzne AWF w Krakowie.
Szopa J. (1986) Genetyczne uwarunkowania psychomotorycznych i fizjologicznych właściwości człowieka. (W:) Motoryczność dzieci i młodzieży – aspekty teoretyczne oraz implikacje metodyczne, (red.) J. Raczek. Cz. I. AWF Katowice.
Szopa J. (1988) W poszukiwaniu struktury motoryczności. Analiza czynnikowa cech somatycznych, funkcjonalnych i prób sprawności fizycznej u chłopców i dziewcząt w wieku 8–19 lat. Wydawnictwo Monograficzne AWF Kraków, nr 35.
Szopa J. (1989a) Nowa koncepcja klasyfikacji i struktury motoryczności człowieka. „Antropomotoryka”, nr 2, s. 3–8.
Szopa J. (1989b) Z badań nad strukturą motoryczności: analiza czynnikowa predyspozycji oraz efektów motorycznych u chłopców i dziewcząt w wieku 18–19 lat. „Antropomotoryka”, nr 2, s. 45–71.
Szopa J. (1992) Zarys antropomotoryki. Wydawnictwo Skryptowe AWF Kraków, nr 117.
Szopa J. (1993) Raz jeszcze o strukturze motoryczności – próba syntezy. „Antropomotoryka”, nr 10, s. 217–227.
Szopa J., Wątroba J. (1992) Dalsze badania nad strukturą motoryczności ze szczególnym uwzględnieniem uzdolnień ruchowych. „Antropomotoryka”, nr 8, s. 3–37.
Szopa J., Chwała W., Ruchlewicz T. (1999) Identification structure and validity of technic of motor abilities. „Journal of Human Kinetics”, vol. I, „Antropomotoryka”, nr 19–20, Kraków, s. 5–23.
Szopa J., Mleczko E., Cempla J. (1985) Zmienność oraz genetyczne i środowiskowe uwarunkowania podstawowych cech psychomotorycznych i fizjologicznych w populacji wielkomiejskiej, w przedziale wieku 7–62 lat. Monografia nr 25, AWF Kraków.
Szopa J., Mleczko E., Żak S. (1996) Podstawy antropomotoryki. PWN, Warszawa–Kraków.
Ślężyński J., Mynarski W. (1973) Współzależności cech morfologicznych, fizjologicznych i motorycznych u mężczyzn w wieku 19–46 lat. „Kultura Fizyczna”, nr 8, s. 344–348.
Tanner J.M. (1963) Rozwój w okresie pokwitania. PZWL, Warszawa.
Trzaskoma Z., Trzaskoma Ł. (2001) Kompleksowe zwiększanie siły mięśniowej sportowców. Centralny Ośrodek Sportu, Warszawa.
Trześniowski R. (1982) Zagadnienie sprawności fizycznej młodzieży polskiej w pracach naukowych AWF. Międzynarodowa Konferencja Naukowa, AWF Warszawa.
Turewski I.M. (1993) Struktura psichofiziczeskoj podgotowlennosti uczaszczejsia molodeżi. Ministerstwo Obrazowanija Rosijskoj Federacii, Tuła.
Ulatowski T. (1981) Teoria i metodyka sportu. Sport i Turystyka, Warszawa.
Ulmer H.V., Gabriel O. (1997) Testspezitat bei der Motordiagnostik – Ein kritischservergleich von Abałakow – und Jump-and-reach-Test. Vortrag für Kongres „Sport Kinetics ’97”, Magdeburg.
Verchośanskij Ju.V., Dobrovolskij I.M., Śuplecov S.N., Ćuregin A.I., Reva V.K. (1977) Faktornaja struktura skorostno-silovych kaćestv u detiej. „Teorija i Praktika Fizi eskoj Kultury”, 1, s. 40–43.
Viitasalo J.T., Komi P.V. (1981) Effects of fatigue on isometric force – and relaxation-time characteristics in human muscles. „Europ. J. Appl. Physiol.”, 49, s. 7–16.
Vilkner H.J. (1986) Zur Vervollkommung der motorischen Reaktionsfähigkeit im Sport Unterricht. „Körpererziehung“, Nr. 5, s. 181–191.
Vilkner H.J. (1989) Ansätze zur Charakteristik der Beziehungen zwissen Koordinativen und Konditionellen Fähigkeiten. „Theor. Prax. Körperkult.“, Beiheft 2, s. 112–113.
Wachowski E. (1981) Współzależność cech motorycznych w procesie szkolenia sportowego. I Kongres Naukowy Kultury Fizycznej. Materiały i dokumenty. GKKFiS, Warszawa, s. 357–360.
Wachowski E., Strzelczyk R. (1991) Atuty cech motorycznych. „Trening”, nr 1, s. 106–114.
Wachowski E., Strzelczyk R. (1999) Trafność pomiaru motorycznych cech kondycyjnych. Monografie nr 342, AWF Poznań.
Wachowski E., Osiński W., Winkler A. (1976) Zależność między siłą mięśniową i mocą a poziomem maksymalnej prędkości biegowej. „Kultura Fizyczna”, nr 6, s. 254–257.
Wachowski E., Strzelczyk R., Osiński W. (1987) Pomiar cech sprawności motorycznej osobników uprawiających sport. Wyniki badań. Monografie nr 238, AWF Poznań, s. 59–60.
Waśkiewicz Z. (2002) Wpływ wysiłków anaerobowych na wybrane aspekty koordynacji motorycznej. AWF Katowice.
Ważny Z. (1977) Trening siły mięśniowej. Sport i Turystyka, Warszawa.
Wilimczik K., Roth K. (1983) Bewegungslehre. Rowohlt, Hamburg.
Wilkie D.R. (1974) Mięsień. PZWL, Warszawa.
Wiszniakow A.W. (1993). Struktura koordinacionnych sposobnostiej i metodika ich kontrolja u detej 11–12 lat. (Dissertacja kandidata pedagogiczeskich nauk). Moskwa.
Wit A. (1980) Zagadnienia regulacji w procesie rozwoju siły mięśniowej na przykładzie zawodników uprawiających podnoszenie ciężarów. Instytut Sportu, Warszawa.
Wolański N. (1981) Genetyczne i antropologiczne czynniki osiągnięć sportowych i rozwoju motorycznego. „Kultura Fizyczna”, nr 2: s. 1–7, nr 3: s. 1–8.
Wolański N. (1983) Rozwój biologiczny człowieka. Cz. 1, 2. PWN, Warszawa, ss. 705.
Wolański N., Kasprzak E. (1979) Similarity in some physiological, biochemical and psychomotor traits between parents and 2–45 years old offspring. „Studies in Human Ecology”, vol. 3.
Wolański N., Parižkowa J. (1976) Sprawność fizyczna a rozwój człowieka. Sport i Turystyka, Warszawa.
Wyżnikiewicz-Kopp Z. (1986) Sport w rozwoju i wychowaniu dzieci i młodzieży (Wychowanie, trening, selekcja). Uniwersytet Szczeciński, Szczecin.
Wyżnikiewicz-Kopp Z. (1992) Koordynacyjne zdolności ruchowe dzieci i młodzieży. Rozprawy i Studia nr 23, Uniwersytet Szczeciński, Szczecin.
Yandell K.M., Spirduso W.M. (1981) Sex and athletic status as factors in reaction latency and movement time. „Res. Quart. Exerc. and Sport”, 52, 4.
Zaciorski W.M. (1969) Zależności między cechami motorycznymi. „Sport Wyczynowy”, nr 10, s. 7–16.
Zaciorski W.M. (1970) Kształcenie cech motorycznych człowieka. „Sport i Turystyka”, Warszawa.
Zaciorski W.M. (1979) Osnowy sportiwnoj metrologii. Fizkultura i Sport, Moskwa.
Zaciorski W., Kulig N.G., Smirnov J. (1970) Zależność między cechami motorycznymi. „Sport Wyczynowy”, Cz. I–10:7-13. Cz. II i III–1:25-28; 2:1-4.
Zaciorski W.M. i in. (1978) Czynniki wpływające na celność strzałów piłkarza. „Sport Wyczynowy”, nr 5, s. 37.
Zimmermann K. (1982) Wesentliche koordinative Fahigkeiten fur Sportspiele. „Theor. Prax. Körperkult.”, Nr. 6.
Zimmermann K. (1983) Zur Weiterentwicklung der Theorie der koordinativen Fähigkeiten. „Wissenschaftliche Zeitschrift der DHfK“, 3, Leipzig, s. 33–44.
Zimmermann K. (1986) Auspragung koordinativen Fahigkeiten im Handballtraining. „Körpererziehung“, Nr. 5.
Zimmermann K., Nicklisch R. (1981) Die Ausbildung koordinativen Fähigkeiten und ihre Bedeutung für die technische bzw. technisch-taktische Leistungsfähigkeiten der sportler. „Theor. Prax. Körperkult.“, Nr. 10.
The structure of motor capabilities and their global and local forms
The problem of the relation between global and local motor capabilities has not been taken up in research. There has also been a scarcity of complex solutions to the problem of relation between conditional and coordination motor capabilities. What’s more, there have been few researches into precise evincing of various motor capabilities. Considering the importance of the problem an attempt at finding its solution has been take up. That is the reason for having tested global and local evincing if conditional and coordination motor capabilities, which aims at:
Determining the relation between local and global evincing of conditional and coordination motor capabilities.
Recognizing the relation between conditional and coordination motor capabilities.
Determining the relation between an optimum precise and maximal evincing of motor capabilities in conditional and coordination exercises of global and local character as well as ascertaining the level of accuracy of the motor exercise taking.
Establishing a level of symmetrization of global and local evincing of training and coordination motor capabilities.
Searching for sexual dimorphism on the subject of global and local evincing of conditional and coordination motor capabilities.
The research has been carried out on a group of 176 male students and, in order to estimate sexual dimorphism, the results obtained by 84 female students have been included. Of all the values involved in the measurements 25 most representative have been chosen and then divided into the four following groups:
global conditional motor capabilities
local conditional motor capabilities
global coordination motor capabilities
local coordination motor capabilities
The exercises involving numerous muscular and joints group to be performed properly have been regarded as global, and those with movement involving a single basic muscular group activity within one joint – as local exercises.
The selecting of testing exercises aiming at estimating the level of conditional motor capabilities has been based on their usefulness in measurement of maximal strength, power and velocity evincing in its elementary forms, such as: response time, a single movement velocity and movement frequency. In order to estimate coordination motor capabilities there have been used such motor exercises that require rapid response and either time or time – space accuracy in single and cyclic movements.
The maximal motor test results have provided the base to estimate accuracy measure for each tested individual (in percentages) which enabled us to compare deviations from a set value of various motor capabilities measured in various units. Deviation values determined the measure of particular kind of accuracy. The potential for evincing global and local motor capabilities were compared in males and females using a dimorphism indicator. A similar procedure was followed to estimate a symmetrization indicator, which is a measure of similarity of motor capabilities evincing in activities done with a right or left limb or turning right or left.
The obtained results were statistically elaborated with a “STATISTICA 6,0 PL” program. Arithmetic means and significance of differences between them were calculated. Besides, the calculations included correlation matrixes between the analysed motor capabilities and multiple regressions by means of a method of reverse stepwise regression. In order to estimate similarities between the analysed variables they were divided into groups using characteristic aglomeration.
The results of the analysis let us ascertain that:
Local and global coordination motor capabilities were irrelevant. Only within local and global training capabilities evincing was there partial relevance. That means developing coordination capabilities requires using adequate training exercise. However, global and local training motor capabilities can lack some specifity which should make it easier to select suitable means used in their development.
There is no foundation for claiming there are strict coordination conditions for training motor capabilities regardless of global and local character of motor capabilities evincing. Such conclusion suggests using physical exercise involving informative and energy processes in development of both groups and motor capabilities.
An appropriate proportioning of the selected motor capabilities depended on their maximal value. Besides, evincing of the appropriateness in various motor exercises was conditioned by the capability level of activity kinesthetic differentation and complexity level of an physical exercise. The precision, which is, a precise performing of an exercise depends on the possibility of arranging its performance as well as monitoring and correcting the plan accomplishment. However, local and global character of the physical exercise is of little importance.
The result of performing most of the analysed exercises requiring training and coordination motor capabilities to be evinced were characterized by a considerable degree of symmetry with the exception of local strength and movements frequency appropriateness, which were both considerably asymmetric. The differences in evincing motor capabilities in both upper limbs are likely to have resulted from the complex role they play in daily routines, which means, they were more of an acquired character rather than congenital one. The lateralization conditioned by the nervous system structure seems to evince less considerably in local motor exercises requiring higher precision. Restricted possibilities of symmetric performing of motor exercises might be higher in case of maximal activity rather than optimum one.
The results acquired by the females and males were alike within the range of the inclined local and global motor capabilities conditioned by the strength and mobility of the nervous system. However, the differentiation in the level of motor capabilities both in females and males determined by their position in the structure, dividing them into training and coordination ones, suggests they need to be shaped individually for each sex.The problem taken up in the research requires further observations based on the research material which varies in age, body build, sex and trained discipline (occupation) as well as in sports advancement (job proficiency).
STRUKTUR DER MOTORISCHEN FÄHIGKEITEN UND SEINE GLOBALES UND LOKALES ERSCHEINEN
Die Problematik der Abhängigkeiten zwischen globalen und lokalen motorischen Fähigkeiten wurde nicht allzu oft in Untersuchungen aufgenommen. Es fehlten auch komplexe Lösungen, die die Relationen zwischen globalen und lokalen motorischen Fähigkeiten betreffen. Es gab auch keine vielen Untersuchungen zum genauen Erscheinen verschiedener motorischen Fähigkeiten. Wegen der Gewichtigkeit dieser Probleme wurde der Versuch unternommen, sie komplex zu lösen. Deswegen wurden globales und lokales Erscheinen der konditionellen und der koordinierenden motorischen Fähigkeiten untersucht, mit dem Bestreben folgende Ziele zu erreichen:
Bestimmung der Abhängigkeiten zwischen globalem und lokalem Erscheinen von konditionellen und koordinierenden motorischen Fähigkeiten.
Erforschung der Verhältnisse zwischen konditionellen und koordinierenden motorischen Fähigkeiten.
Bestimmung der Verbindungen zwischen optimalem (genauem) und maximalem Erscheinen der motorischen Fähigkeiten bei konditionellen und koordinierenden Aufgaben mit globalem und lokalem Charakter und Feststellung von Genauigkeit der Erfüllung der motorischen Aufgaben.
Bestimmung des Grades von Umsymmetrierung des lokalen und des globalen Erscheinens von konditionellen und koordinierenden motorischen Fähigkeiten.
Suche nach Anzeichen vom Geschlechtsdimorphismus im Bereich von globalem und lokalem Erscheinen der konditionellen und koordinierenden motorischen Fähigkeiten.
Die Untersuchung hat eine Gruppe von 176 Studenten umfasst und zur Beurteilung vom Geschlechtsdimorphismus wurden die Ergebnisse von 84 Studentinnen verwendet. Von allen bei der Erhebung berücksichtigten Indikatoren wurden 25 repräsentativste ausgesucht und in vier folgende Gruppen geteilt:
globale konditionelle motorische Fähigkeiten;
lokale konditionelle motorische Fähigkeiten;
globale koordinierende motorische Fähigkeiten;
lokale koordinierende motorische Fähigkeiten.
Als globale wurden solche Aufgaben erachtet, bei denen der Einsatz der mehreren Muskelgruppen und Gelenke für ihre richtige Ausübung notwendig war. Zu den lokalen wurden diese gerechnet, bei denen die Bewegung einer Aktivität von einer grundlegenden Muskelgruppe bedarf und im Bereich eines Gelenkes verlief.
Die Auswahl der Testaufgaben zur Beurteilung des Niveaus von konditionellen motorischen Fähigkeiten wurde anhand der Verwendbarkeit der maximalen Stärke, Kraft und Schnelligkeit, die in den elementaren Formen, d.h. Zeit der Reaktion, Geschwindigkeit der einzelnen Bewegung und der Häufigkeit der Bewegungen erscheint, ausgesucht. Zur Beurteilung der koordinierenden motorischen Fähigkeiten wurden solchen motorischen Aufgaben verwendet, bei denen schnelle Reaktion und zeitliche Genauigkeit oder zeitlich-räumliche Reaktion bei den einzelnen oder zyklischen Bewegungen notwendig waren.
Aufgrund der maximalen Ergebnisse der Erfüllung der motorischen Aufgaben wurde das Maß der Genauigkeit in Prozenten für jeden Untersuchten errechnet. Deswegen hatte man die Möglichkeit gewonnen, die Abweichungen vom Zielwert für verschiedene motorische Fähigkeiten, in verschiedenen Einheiten gemessen, zu vergleichen. Die Abweichungswerte wurden als Maß einzelner Arten der Genauigkeit erachtet. Die Möglichkeiten des Erscheinens von globalen und lokalen motorischen Fähigkeiten bei Frauen und Männern wurden anhand des Indikators des Dimorphismus verglichen.
Ähnliche Vorgehensweise wurde zur Bestimmung des Indikators von Umsymmetrierung angenommen, die das Maß der Ähnlichkeit von Anzeichen der motorischen Fähigkeiten bei den mit der rechten und der linken Hand oder in die rechte und die linke Seite ausgeübten Bewegungen.
Die gewonnen Ergebnisse wurden statistisch mit dem Programm “STATISTICA 6,0 PL“ bearbeitet. Es wurden arithmetische Durchschnitte und die Wesentlichkeit der Differenzen zwischen ihnen berechnet. Es wurden auch Matrix der Korrelation zwischen den analysierten motorischen Fähigkeiten und mehrfache Regression anhand der Methode der Rückschrittregression berechnet. Wegen der Bestimmung der Ähnlichkeiten zwischen den analysierten Variablen wurde eine Gruppierung mit der Verwendung der Agglomeration der Merkmale unternommen.
In Folge der durchgeführten Analyse wurde es bestimmt:
Lokale und globale koordinierende motorische Fähigkeiten waren unabhängig. Die Verbindungen sind teilweise nur im Bereich der lokalen und globalen Anzeichen der konditionellen motorischen Fähigkeiten aufgetreten. Dies bedeutet Bedarf der Entwicklung der koordinierenden Fähigkeiten mit dem Einsatz der für sie spezifischen Trenningsmittel. Aber globale und lokale konditionelle motorische Fähigkeiten können sich durch einen gewissen Mangel an Spezifischem auszeichnen, was die Auswahl der eingesetzten Mittel für ihre Entwicklung vereinfachen sollte.
Es wurden keine Anhaltspunkte für die Schlussfolgerung gefunden, dass starke koordinierende Bedingungen der konditionellen motorischen Fähigkeiten bestehen, unabhängig von globalem und lokalem Charakter der Erscheinung von motorischen Fähigkeiten. Im Prozess der Entwicklung der beiden Gruppen der motorischen Fähigkeiten soll man also Bewegungsaufgaben einsetzen, die die Mitwirkung der Informations- und Energieprozesse notwendig machen.
Genaue (Dosierung) Wiedergabe der aufgegebenen Größe der ausgesuchten motorischen Fähigkeiten war von ihrem maximalen Wert abhängig. Außerdem war das Erscheinen der Genauigkeit bei verschiedenen motorischen Aufgaben vom Niveau der kinestetischen Fähigkeit der Differenzierung der Bewegung und vom Schwierigkeitsgrad der Bewegungsaufgabe bedingt. Die Präzision also die genaue Ausführung der Bewegung ist von der Möglichkeit der Planung ihrer Ausführung und der Kontrolle und von der Art der Korrektur der Planausführung abhängig. Aber lokaler oder globaler Charakter der Bewegungsaufgaben hat keine besondere Bedeutung.
Die Ergebnisse der Ausführung der Mehrheit von analysierten Aufgaben, die die Aufweisung der konditionellen und koordinierenden motorischen Fähigkeiten notwendig machten, kennzeichnete ein bedeutender Grad der Symmetrie. Die Ausnahme bildete die lokale Genauigkeit der Kraft und die Genauigkeit der lokalen Häufigkeit der Bewegungen, die eindeutig asymmetrisch waren. Höchstwahrscheinlich ergaben sich die Differenzen bei der Erscheinung der motorischen Fähigkeiten der beiden oberen Gliedmassen aus der unterschiedlichen Rolle bei der Ausführung der Lebenstätigkeiten, also hatten sie eher einen erlernten Charakter als einen angeborenen. Es scheint, dass die Lateralisierung, die vom Bau des Nervensystems abhängig ist, im geringeren Masse bei den lokalen motorischen Aufgaben erscheint, die mehr Genauigkeit bedürfen. Man kann vermuten, dass die Beschränkungen im Bereich der Möglichkeiten der Symmetrie der Ausführung der motorischen Aufgaben größer im Falle der Aufgaben mit der größeren als optimaler Aktivität sind.
Frauen und Männer haben ähnliche Ergebnisse im Bereich der bei den Untersuchungen berücksichtigten, lokalen und globalen motorischen Fähigkeiten erreicht, die von Kraft und Bewegung bedingt sind. Aber die Differenzierung des Niveaus der motorischen Fähigkeiten bei Frauen und Männern in Abhängigkeit von ihrer Stellung in der Struktur, die sie zu konditionellen und koordinierenden teilt, weist auf den Bedarf der einzelnen Entwicklung bei den Vertretern beider Geschlechter hin. Die in der Arbeit aufgenommene Problematik bedarf weiterer Beobachtung aufgrund des Untersuchungsstoffes, der in Hinsicht auf Alter, Körperbau, Geschlecht und Art der ausgeübten Sportdisziplin (des ausgeübten Berufes) und des sportlichen Fortschritts (Berufsfertigkeit) differenziert wird.